

Role of JC virus agnoprotein in DNA repair
- PMID: 15280468
- PMCID: PMC479055
- DOI: 10.1128/JVI.78.16.8593-8600.2004
Role of JC virus agnoprotein in DNA repair
Abstract
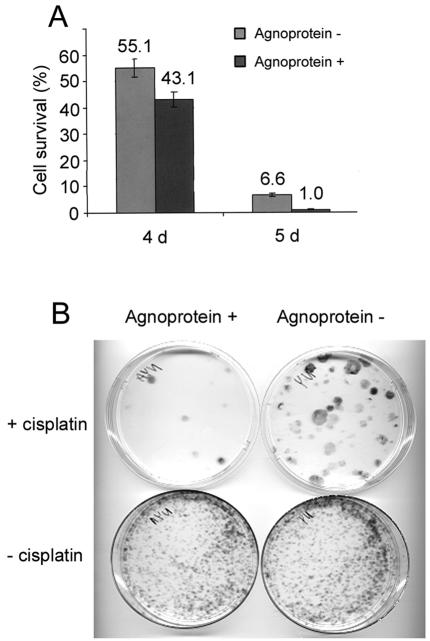
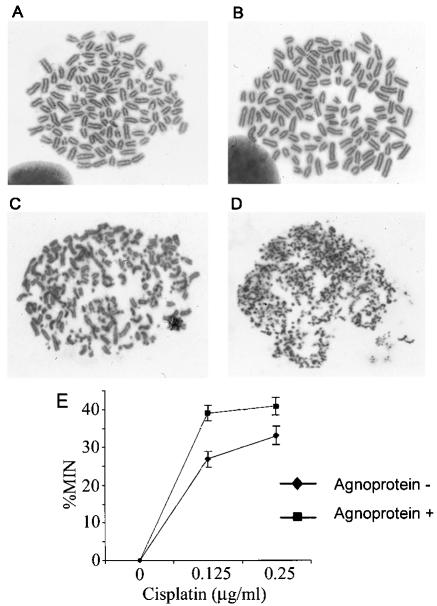
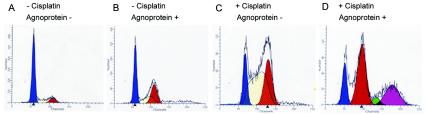
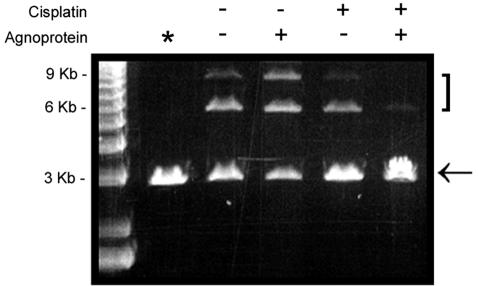
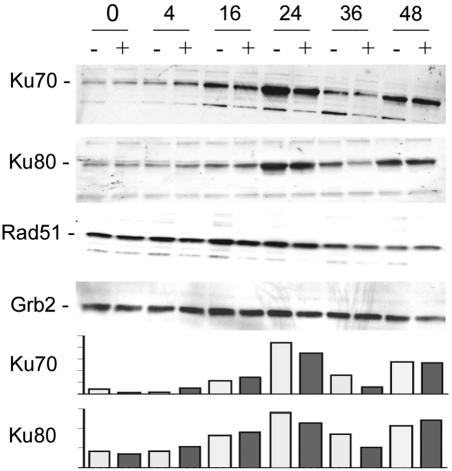
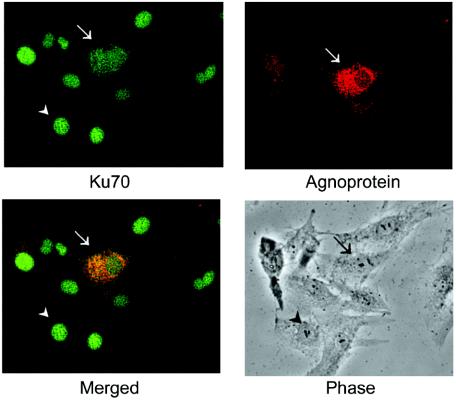
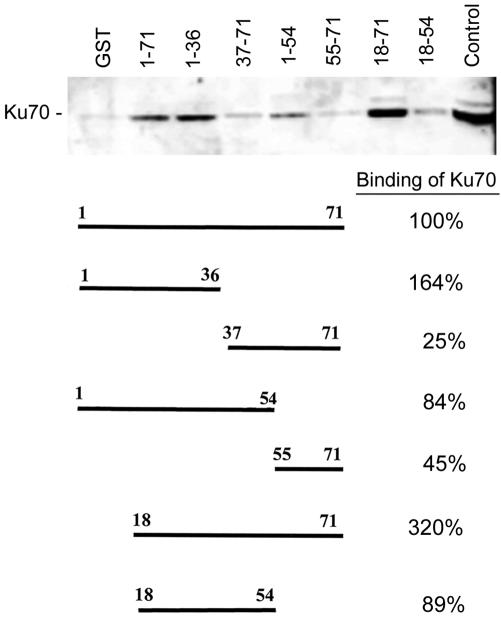
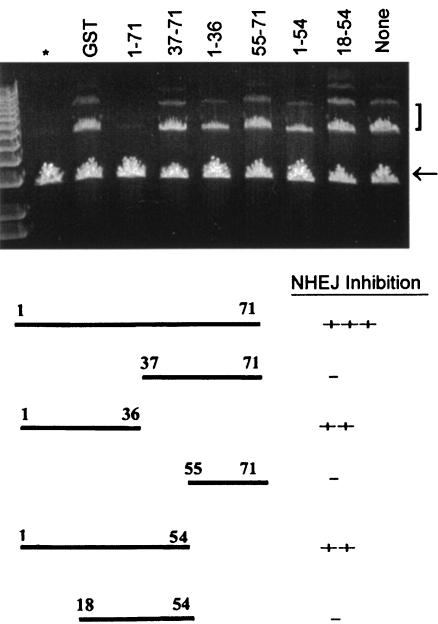
The late region of human neurotropic JC virus encodes a small 71-amino-acid agnoprotein that is also found in the polyomaviruses simian virus 40 and BK virus. Several functions of agnoprotein have been identified, including roles in regulating viral transcription and virion maturation. Earlier studies showed that agnoprotein expressed alone induced p21/WAF-1 expression and caused cells to accumulate in the G(2)/M stage of the cell cycle. Here we report that agnoprotein expression sensitized cells to the cytotoxic effects of the DNA-damaging agent cisplatin. Agnoprotein reduced the viability of cisplatin-treated cells and increased chromosome fragmentation and micronucleus formation. Whereas cisplatin-treated control cells accumulated in S phase, cells expressing agnoprotein did not, instead becoming aneuploid. Agnoprotein expression correlated with impaired double-strand-break repair activity in cellular extracts and reduced expression of the Ku70 and Ku80 DNA repair proteins. After agnoprotein expression, much of the Ku70 protein was located in the perinuclear space, where agnoprotein was also found. Results from binding studies showed an interaction of agnoprotein with Ku70 which was mediated by the N terminus. The ability of agnoprotein to inhibit double-strand break repair activity when it was added to cellular extracts was also mediated by the N terminus. We conclude that agnoprotein inhibits DNA repair after DNA damage and interferes with DNA damage-induced cell cycle regulation. Since Ku70 is a subunit of the DNA-dependent protein kinase that is responsible both for double-strand break repair and for signaling damage-induced cell cycle arrest, the modulation of Ku70 and/or Ku80 by agnoprotein may represent an important event in the polyomavirus life cycle and in cell transformation.
Figures
References
-
- Berger, J. R. 2003. Progressive multifocal leukoencephalopathy in acquired immuno-deficiency syndrome: explaining the high incidence and disproportionate frequency of the illness relative to other immunosuppressive conditions. J. Neurovirol. 9(Suppl. 1):38-41. - PubMed
-
- Berger, J. R., and M. Concha. 1995. Progressive multifocal leukoencephalopathy: the evolution of a disease once considered rare. J. Neurovirol. 1:5-18. - PubMed
-
- Cole, C. N. 1996. Polyomavirinae: the viruses and their replication, p. 917-946. In B. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fundamental virology, 3rd ed. Lippincott Williams & Wilkins, Philadelphia, Pa.
-
- Cromie, G. A., J. C. Connelly, and D. R. Leach. 2001. Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans. Mol. Cell 8:1163-1174. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
