

Mrc1 is required for sister chromatid cohesion to aid in recombination repair of spontaneous damage
- PMID: 15282308
- PMCID: PMC479732
- DOI: 10.1128/MCB.24.16.7082-7090.2004
Mrc1 is required for sister chromatid cohesion to aid in recombination repair of spontaneous damage
Abstract
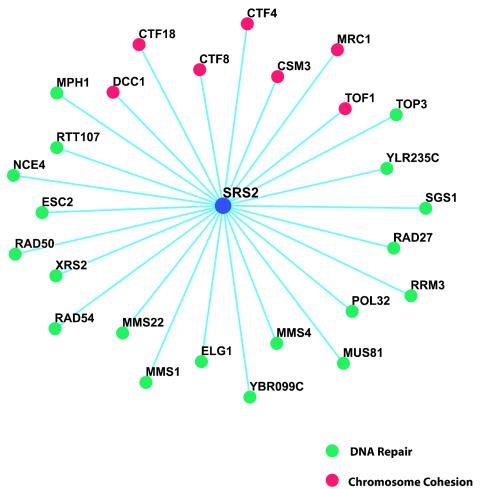
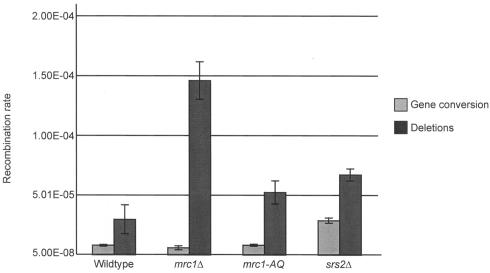
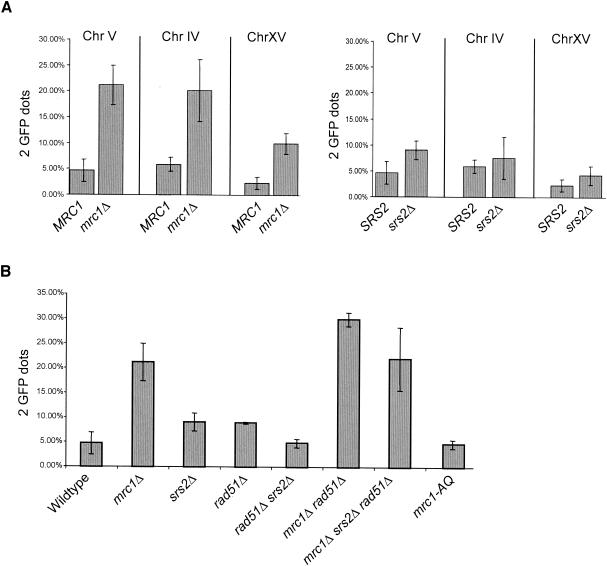
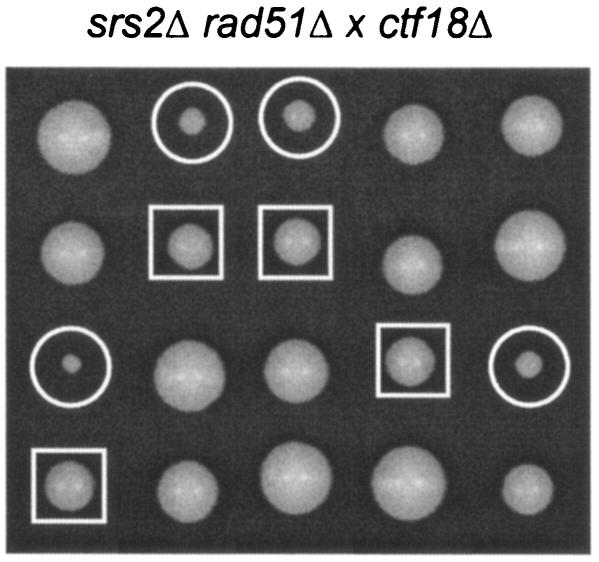
The SRS2 gene of Saccharomyces cerevisiae encoding a 3'-->5' DNA helicase is part of the postreplication repair pathway and functions to ensure proper repair of DNA damage arising during DNA replication through pathways that do not involve homologous recombination. Through a synthetic gene array analysis, genes that are essential when Srs2 is absent have been identified. Among these are MRC1, TOF1, and CSM3, which mediate the intra-S checkpoint response. srs2 Delta mrc1 Delta synthetic lethality is due to inappropriate recombination, as the lethality can be suppressed by genetic elimination of homologous recombination. srs2 Delta mrc1 Delta synthetic lethality is dependent on the role of Mrc1 in DNA replication but independent of the role of Mrc1 in a DNA damage checkpoint response. mrc1 Delta, tof1 Delta and csm3 Delta mutants have sister chromatid cohesion defects, implicating sister chromatid cohesion established at the replication fork as an important factor in promoting repair of stalled replication forks through gap repair.
Figures



References
-
- Alcasabas, A. A., A. J. Osborn, J. Bachant, F. Hu, P. J. Werler, K. Bousset, K. Furuya, J. F. Diffley, A. M. Carr, and S. J. Elledge. 2001. Mrc1 transduces signals of DNA replication stress to activate Rad53. Nat. Cell Biol. 3:958-965. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous