

Inhibition of nuclear import by the proapoptotic protein CC3
- PMID: 15282309
- PMCID: PMC479715
- DOI: 10.1128/MCB.24.16.7091-7101.2004
Inhibition of nuclear import by the proapoptotic protein CC3
Abstract
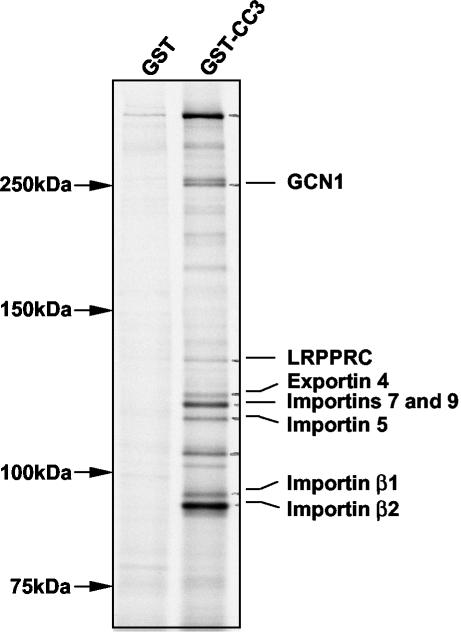
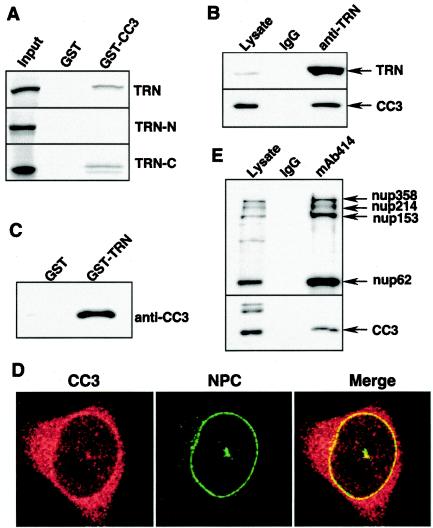
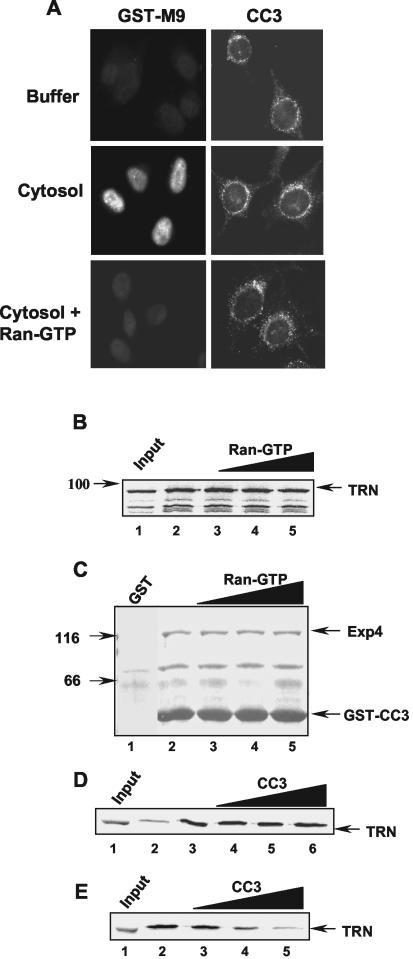
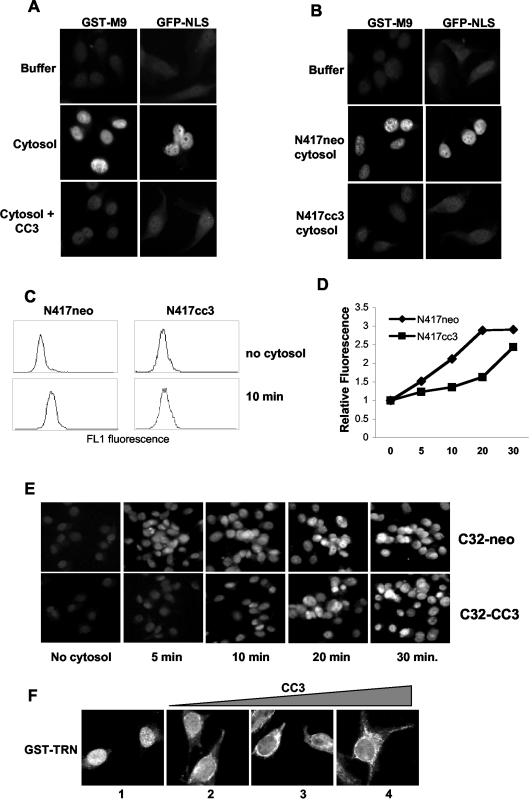
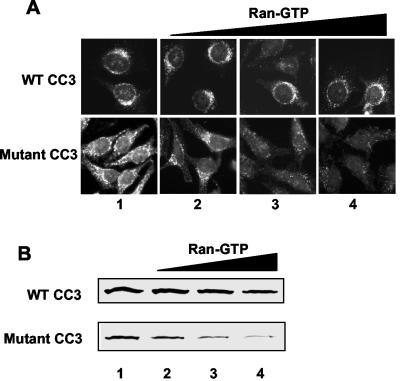
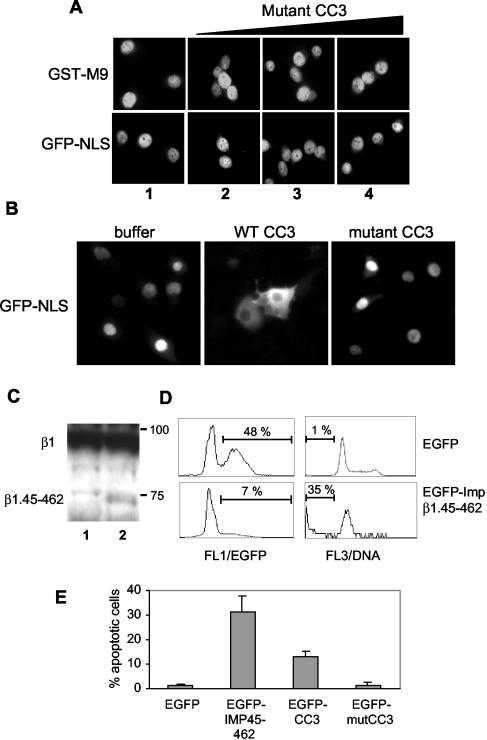
We report here that the normal cellular protein CC3/TIP30, when in excess, inhibits nuclear import in vitro and in vivo. CC3 binds directly to the karyopherins of the importin beta family in a RanGTP-insensitive manner and associates with nucleoporins in vivo. CC3 inhibits the nuclear import of proteins possessing either the classical nuclear localization signal or the M9 signal recognized by transportin. CC3 also inhibits nuclear translocation of transportin itself. Cells modified to express higher levels of CC3 have a slower rate of nuclear import and, as described earlier, show an increased sensitivity to death signals. A mutant CC3 protein lacking proapoptotic activity has a lower affinity for transportin, is displaced from it by RanGTP, and fails to inhibit nuclear import in vitro and in vivo. Together, our results support a correlation between the ability of CC3 to form a RanGTP-resistant complex with importins, inhibit nuclear import, and induce apoptosis. Significantly, a dominant-negative form of importin beta1 shown previously to inhibit multiple transport pathways induces rapid cell death, strongly indicating that inhibition of nuclear transport serves as a potent apoptotic signal.
Figures
Similar articles
-
The importin-beta binding domain of snurportin1 is responsible for the Ran- and energy-independent nuclear import of spliceosomal U snRNPs in vitro.J Cell Biol. 2002 Feb 4;156(3):467-79. doi: 10.1083/jcb.200108114. Epub 2002 Jan 28. J Cell Biol. 2002. PMID: 11815630 Free PMC article.
-
The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus.EMBO J. 1997 Nov 3;16(21):6535-47. doi: 10.1093/emboj/16.21.6535. EMBO J. 1997. PMID: 9351834 Free PMC article.
-
Multiple mechanisms promote the inhibition of classical nuclear import upon exposure to severe oxidative stress.Cell Death Differ. 2004 Aug;11(8):862-74. doi: 10.1038/sj.cdd.4401432. Cell Death Differ. 2004. PMID: 15088071
-
Structures of importins.Results Probl Cell Differ. 2002;35:93-113. doi: 10.1007/978-3-540-44603-3_5. Results Probl Cell Differ. 2002. PMID: 11791410 Review. No abstract available.
-
Molecular mechanism of translocation through nuclear pore complexes during nuclear protein import.FEBS Lett. 2001 Jun 8;498(2-3):145-9. doi: 10.1016/s0014-5793(01)02489-9. FEBS Lett. 2001. PMID: 11412846 Review.
Cited by
-
Revisiting Notch in remyelination of multiple sclerosis lesions.J Clin Invest. 2009 Jan;119(1):10-3. doi: 10.1172/JCI37786. J Clin Invest. 2009. PMID: 19104146 Free PMC article.
-
Association of TIP30 expression and prognosis of hepatocellular carcinoma in patients with HBV infection.Cancer Med. 2016 Sep;5(9):2180-9. doi: 10.1002/cam4.728. Epub 2016 Jul 15. Cancer Med. 2016. PMID: 27418384 Free PMC article.
-
Subcongenic analysis of a quantitative trait locus affecting body weight and glucose metabolism in zinc transporter 7 (znt7)-knockout mice.BMC Genet. 2019 Feb 18;20(1):19. doi: 10.1186/s12863-019-0715-2. BMC Genet. 2019. PMID: 30777014 Free PMC article.
-
Mitochondrial protein targets of thiol-reactive electrophiles.Chem Res Toxicol. 2008 Apr;21(4):796-804. doi: 10.1021/tx700433m. Epub 2008 Mar 7. Chem Res Toxicol. 2008. PMID: 18324786 Free PMC article.
-
Karyopherin Alpha Proteins Regulate Oligodendrocyte Differentiation.PLoS One. 2017 Jan 20;12(1):e0170477. doi: 10.1371/journal.pone.0170477. eCollection 2017. PLoS One. 2017. PMID: 28107514 Free PMC article.
References
-
- Adam, S. A., and L. Gerace. 1991. Cytosolic proteins that specifically bind nuclear location signals are receptors for nuclear import. Cell 66:837-847. - PubMed
-
- Baker, M. E. 1999. TIP30, a cofactor for HIV-1 Tat-activated transcription, is homologous to short-chain dehydrogenases/reductases. Curr. Biol. 9:R471. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases