

Phycobilisome diffusion is required for light-state transitions in cyanobacteria
- PMID: 15286286
- PMCID: PMC520783
- DOI: 10.1104/pp.104.046110
Phycobilisome diffusion is required for light-state transitions in cyanobacteria
Abstract
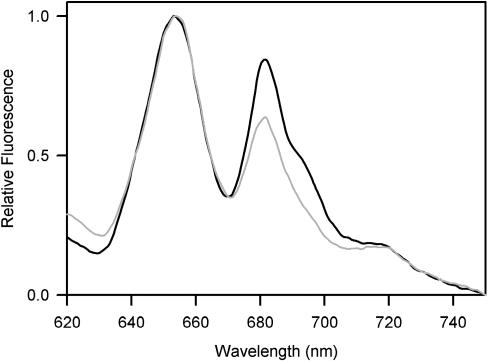
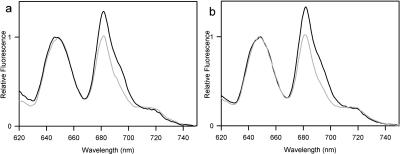
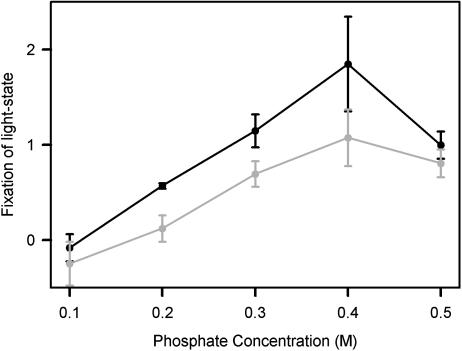
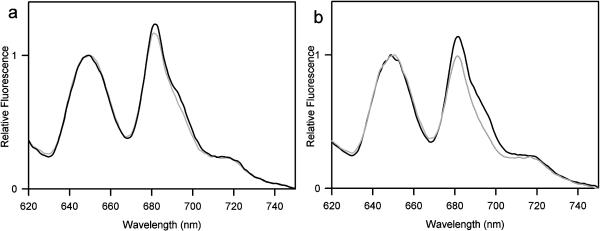
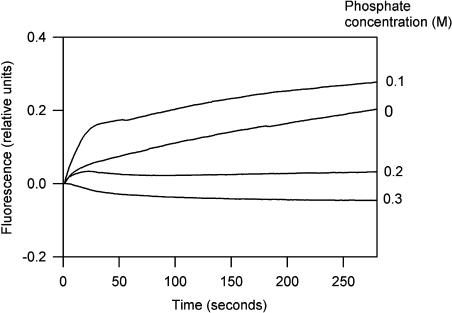
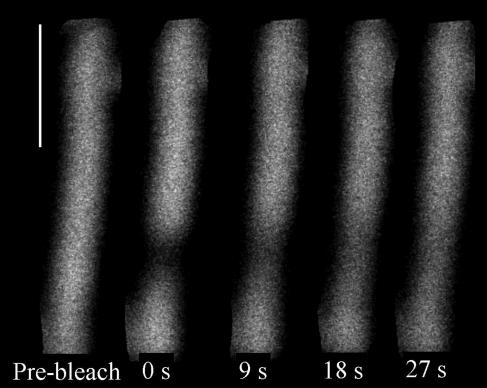
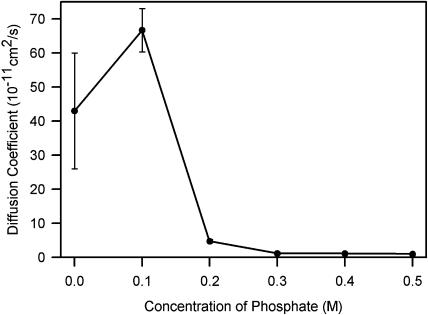
Phycobilisomes are the major accessory light-harvesting complexes of cyanobacteria and red algae. Studies using fluorescence recovery after photobleaching on cyanobacteria in vivo have shown that the phycobilisomes are mobile complexes that rapidly diffuse on the thylakoid membrane surface. By contrast, the PSII core complexes are completely immobile. This indicates that the association of phycobilisomes with reaction centers must be transient and unstable. Here, we show that when cells of the cyanobacterium Synechococcus sp. PCC7942 are immersed in buffers of high osmotic strength, the diffusion coefficient for the phycobilisomes is greatly decreased. This suggests that the interaction between phycobilisomes and reaction centers becomes much less transient under these conditions. We discuss the possible reasons for this. State transitions are a rapid physiological adaptation mechanism that regulates the way in which absorbed light energy is distributed between PSI and PSII. Immersing cells in high osmotic strength buffers inhibits state transitions by locking cells into whichever state they were in prior to addition of the buffer. The effect on state transitions is induced at the same buffer concentrations as the effect on phycobilisome diffusion. This implies that phycobilisome diffusion is required for state transitions. The main physiological role for phycobilisome mobility may be to allow such flexibility in light harvesting.
Figures

Similar articles
-
Distinct roles of CpcG1-phycobilisome and CpcG2-phycobilisome in state transitions in a cyanobacterium Synechocystis sp. PCC 6803.Photosynth Res. 2009 Mar;99(3):217-25. doi: 10.1007/s11120-008-9399-6. Epub 2009 Jan 17. Photosynth Res. 2009. PMID: 19152018
-
Diffusion of phycobilisomes on the thylakoid membranes of the cyanobacterium Synechococcus 7942. Effects of phycobilisome size, temperature, and membrane lipid composition.J Biol Chem. 2001 Dec 14;276(50):46830-4. doi: 10.1074/jbc.M107111200. Epub 2001 Oct 4. J Biol Chem. 2001. PMID: 11590154
-
Phycobilisome Mobility in the Cyanobacterium Synechococcus sp. PCC7942 is Influenced by the Trimerisation of Photosystem I.Photosynth Res. 2004 Feb;79(2):179. doi: 10.1023/B:PRES.0000015399.43503.95. Photosynth Res. 2004. PMID: 16228392
-
Probing the consequences of antenna modification in cyanobacteria.Photosynth Res. 2013 Nov;118(1-2):17-24. doi: 10.1007/s11120-013-9940-0. Epub 2013 Oct 17. Photosynth Res. 2013. PMID: 24132812 Review.
-
Modulating energy arriving at photochemical reaction centers: orange carotenoid protein-related photoprotection and state transitions.Photosynth Res. 2015 Oct;126(1):3-17. doi: 10.1007/s11120-014-0031-7. Epub 2014 Aug 20. Photosynth Res. 2015. PMID: 25139327 Review.
Cited by
-
Cell membrane damage induced by continuous stress of artemisinin sustained-release microspheres (ASMs) on Microcystis aeruginosa at different physiological stages.Environ Sci Pollut Res Int. 2020 Apr;27(11):12624-12634. doi: 10.1007/s11356-020-07813-5. Epub 2020 Jan 31. Environ Sci Pollut Res Int. 2020. PMID: 32006333
-
FRAP analysis on red alga reveals the fluorescence recovery is ascribed to intrinsic photoprocesses of phycobilisomes than large-scale diffusion.PLoS One. 2009;4(4):e5295. doi: 10.1371/journal.pone.0005295. Epub 2009 Apr 20. PLoS One. 2009. PMID: 19381335 Free PMC article.
-
Mechanical regulation of photosynthesis in cyanobacteria.Nat Microbiol. 2020 May;5(5):757-767. doi: 10.1038/s41564-020-0684-2. Epub 2020 Mar 23. Nat Microbiol. 2020. PMID: 32203409
-
Distinct roles of CpcG1-phycobilisome and CpcG2-phycobilisome in state transitions in a cyanobacterium Synechocystis sp. PCC 6803.Photosynth Res. 2009 Mar;99(3):217-25. doi: 10.1007/s11120-008-9399-6. Epub 2009 Jan 17. Photosynth Res. 2009. PMID: 19152018
-
Estimation of relative contribution of "mobile phycobilisome" and "energy spillover" in the light-dark induced state transition in Spirulina platensis.Photosynth Res. 2007 Nov-Dec;94(2-3):315-20. doi: 10.1007/s11120-007-9272-z. Epub 2007 Oct 20. Photosynth Res. 2007. PMID: 17952696
References
-
- Allen JF, Forsberg J (2001) Molecular recognition in thylakoid structure and function. Trends Plant Sci 6: 317–326 - PubMed
-
- Allen JF, Holmes NG (1986) A general model for regulation of photosynthetic unit function by protein phosphorylation. FEBS Lett 202: 175–181
-
- Ashby MK, Mullineaux CW (1999) The role of ApcD and ApcF in energy transfer from phycobilisomes to PSI and PSII in a cyanobacterium. Photosynth Res 61: 169–179
-
- Aspinwall CL, Sarcina M, Mullineaux CW (2004) Phycobilisome mobility in the cyanobacterium Synechococcus sp. PCC7942 is influenced by the trimerisation of Photosystem I. Photosynth Res 79: 179–187 - PubMed
-
- Bruce D, Brimble S, Bryant DA (1989) State transitions in a phycobilisome-less mutant of the cyanobacterium Synechococcus sp. PCC7002. Biochim Biophys Acta 974: 66–73 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
