

The role of the C4 pathway in carbon accumulation and fixation in a marine diatom
- PMID: 15286292
- PMCID: PMC520782
- DOI: 10.1104/pp.104.041319
The role of the C4 pathway in carbon accumulation and fixation in a marine diatom
Abstract
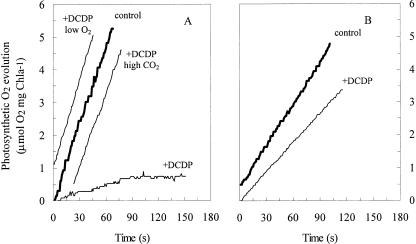
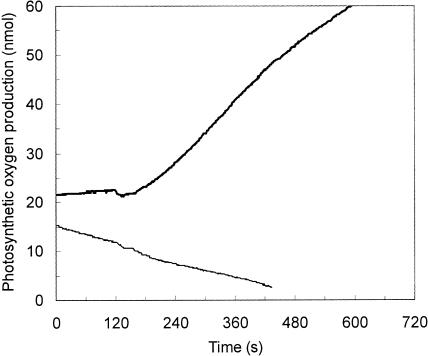
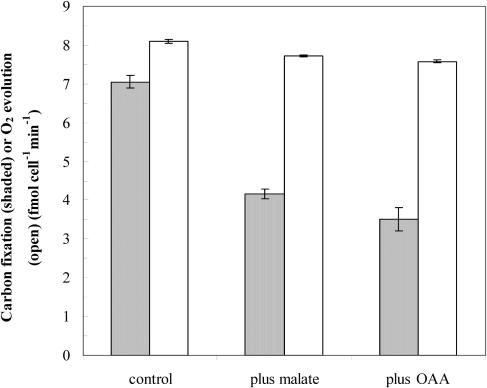
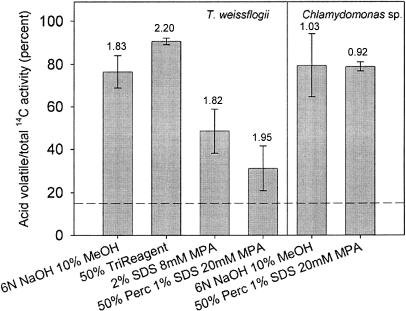
The role of a C(4) pathway in photosynthetic carbon fixation by marine diatoms is presently debated. Previous labeling studies have shown the transfer of photosynthetically fixed carbon through a C(4) pathway and recent genomic data provide evidence for the existence of key enzymes involved in C(4) metabolism. Nonetheless, the importance of the C(4) pathway in photosynthesis has been questioned and this pathway is seen as redundant to the known CO(2) concentrating mechanism of diatoms. Here we show that the inhibition of phosphoenolpyruvate carboxylase (PEPCase) by 3,3-dichloro-2-dihydroxyphosphinoylmethyl-2-propenoate resulted in a more than 90% decrease in whole cell photosynthesis in Thalassiosira weissflogii cells acclimated to low CO(2) (10 microm), but had little effect on photosynthesis in the C(3) marine Chlorophyte, Chlamydomonas sp. In 3,3-dichloro-2-dihydroxyphosphinoylmethyl-2-propenoate-treated T. weissflogii cells, elevated CO(2) (150 microm) or low O(2) (80-180 microm) restored photosynthesis to the control rate linking PEPCase inhibition with CO(2) supply in this diatom. In C(4) organic carbon-inorganic carbon competition experiments, the (12)C-labeled C(4) products of PEPCase, oxaloacetic acid and its reduced form malic acid suppressed the fixation of (14)C-labeled inorganic carbon by 40% to 50%, but had no effect on O(2) evolution in photosynthesizing diatoms. Oxaloacetic acid-dependent O(2) evolution in T. weissflogii was twice as high in cells acclimated to 10 microm rather than 22 microm CO(2), indicating that the use of C(4) compounds for photosynthesis is regulated over the range of CO(2) concentrations observed in marine surface waters. Short-term (14)C uptake (silicone oil centrifugation) and CO(2) release (membrane inlet mass spectrometry) experiments that employed a protein denaturing cell extraction solution containing the PEPCKase inhibitor mercaptopicolinic acid revealed that much of the carbon taken up by diatoms during photosynthesis is stored as organic carbon before being fixed in the Calvin cycle, as expected if the C(4) pathway functions as a CO(2) concentrating mechanism. Together these results demonstrate that the C(4) pathway is important in carbon accumulation and photosynthetic carbon fixation in diatoms at low (atmospheric) CO(2).
Figures
References
-
- Badger MR, Andrews TJ, Whitney SM, Ludwig M, Yellowlees DC, Leggat W, Price GD (1998) The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast-based CO2-concentrating mechanisms in algae. Can J Bot 76: 1052–1071
-
- Beardall J, Mukerji D, Glover HE, Morris I (1976) The path of carbon in photosynthesis by marine phytoplankton. J Phycol 12: 409–417
-
- Bowes G, Rao SK, Estavillo GM, Reiskind JB (2002) C4 mechanisms in aquatic angiosperms: comparisons with terrestrial C4 systems. Funct Plant Biol 29: 379–392 - PubMed
-
-
Burkhardt S, Amoroso G, Riebesell U, Sültemeyer D (2001) CO2 and
uptake in marine diatoms acclimated to different CO2 concentrations. Limnol Oceanogr 46: 1378–1391
-
Burkhardt S, Amoroso G, Riebesell U, Sültemeyer D (2001) CO2 and
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
