

A contribution of mouse dendritic cell-derived IL-2 for NK cell activation
- PMID: 15289500
- PMCID: PMC2211974
- DOI: 10.1084/jem.20040370
A contribution of mouse dendritic cell-derived IL-2 for NK cell activation
Abstract
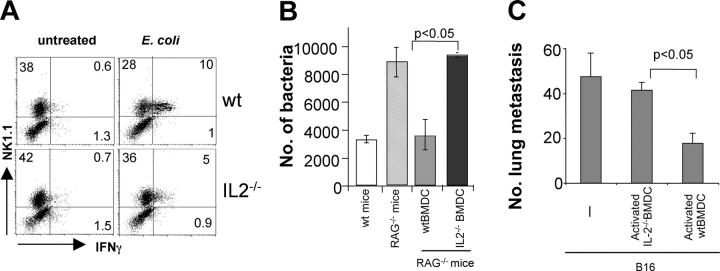
Dendritic cells (DCs) play a predominant role in activation of natural killer (NK) cells that exert their functions against pathogen-infected and tumor cells. Here, we used a murine model to investigate the molecular mechanisms responsible for this process. Two soluble molecules produced by bacterially activated myeloid DCs are required for optimal priming of NK cells. Type I interferons (IFNs) promote the cytotoxic functions of NK cells. IL-2 is necessary both in vitro and in vivo for the efficient production of IFNgamma, which has an important antimetastatic and antibacterial function. These findings provide new information about the mechanisms that mediate DC-NK cell interactions and define a novel and fundamental role for IL-2 in innate immunity.
Figures




Similar articles
-
Rapid generation of potent and tumor-specific cytotoxic T lymphocytes by interleukin 18 using dendritic cells and natural killer cells.Cancer Res. 2000 Sep 1;60(17):4838-44. Cancer Res. 2000. PMID: 10987295
-
Enhancement of human cord blood CD34+ cell-derived NK cell cytotoxicity by dendritic cells.J Immunol. 2001 Feb 1;166(3):1590-600. doi: 10.4049/jimmunol.166.3.1590. J Immunol. 2001. PMID: 11160200
-
IL-4 confers NK stimulatory capacity to murine dendritic cells: a signaling pathway involving KARAP/DAP12-triggering receptor expressed on myeloid cell 2 molecules.J Immunol. 2004 May 15;172(10):5957-66. doi: 10.4049/jimmunol.172.10.5957. J Immunol. 2004. PMID: 15128777
-
The bidirectional crosstalk between human dendritic cells and natural killer cells.J Innate Immun. 2011;3(3):258-63. doi: 10.1159/000323923. Epub 2011 Mar 11. J Innate Immun. 2011. PMID: 21411969 Review.
-
Natural killer-dendritic cell cross-talk in cancer immunotherapy.Expert Opin Biol Ther. 2005 Oct;5(10):1303-15. doi: 10.1517/14712598.5.10.1303. Expert Opin Biol Ther. 2005. PMID: 16197336 Review.
Cited by
-
Inflammatory arthritis can be reined in by CpG-induced DC-NK cell cross talk.J Exp Med. 2007 Aug 6;204(8):1911-22. doi: 10.1084/jem.20070285. Epub 2007 Jul 23. J Exp Med. 2007. PMID: 17646407 Free PMC article.
-
Insights into dendritic cell maturation during infection with application of advanced imaging techniques.Front Cell Infect Microbiol. 2023 Mar 2;13:1140765. doi: 10.3389/fcimb.2023.1140765. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 36936763 Free PMC article. Review.
-
Role of CD14 in host protection against infections and in metabolism regulation.Front Cell Infect Microbiol. 2013 Jul 24;3:32. doi: 10.3389/fcimb.2013.00032. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 23898465 Free PMC article. Review.
-
Rickettsiae stimulate dendritic cells through toll-like receptor 4, leading to enhanced NK cell activation in vivo.J Infect Dis. 2009 Jan 15;199(2):236-42. doi: 10.1086/595833. J Infect Dis. 2009. PMID: 19072551 Free PMC article.
-
NFATc2 mediates epigenetic modification of dendritic cell cytokine and chemokine responses to dectin-1 stimulation.Nucleic Acids Res. 2015 Jan;43(2):836-47. doi: 10.1093/nar/gku1369. Epub 2014 Dec 30. Nucleic Acids Res. 2015. PMID: 25550437 Free PMC article.
References
-
- Granucci, F., C. Vizzardelli, N. Pavelka, S. Feau, M. Persico, E. Virzi, M. Rescigno, G. Moro, and P. Ricciardi-Castagnoli. 2001. Inducible IL-2 production by dendritic cells revealed by global gene expression analysis. Nat. Immunol. 2:882–888. - PubMed
-
- Ricciardi-Castagnoli, P., and F. Granucci. 2002. Opinion: interpretation of the complexity of innate immune responses by functional genomics. Nat. Rev. Immunol. 2:881–889. - PubMed
-
- Huang, Q., D. Liu, P. Majewski, L.C. Schulte, J.M. Korn, R.A. Young, E.S. Lander, and N. Hacohen. 2001. The plasticity of dendritic cell responses to pathogens and their components. Science. 294:870–875. - PubMed
-
- Guermonprez, P., J. Valladeau, L. Zitvogel, C. Thery, and S. Amigorena. 2002. Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 20:621–667. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
