

Characterization of a novel plasma membrane protein, expressed in the midgut epithelia of Bombyx mori, that binds to Cry1A toxins
- PMID: 15294792
- PMCID: PMC492382
- DOI: 10.1128/AEM.70.8.4604-4612.2004
Characterization of a novel plasma membrane protein, expressed in the midgut epithelia of Bombyx mori, that binds to Cry1A toxins
Abstract
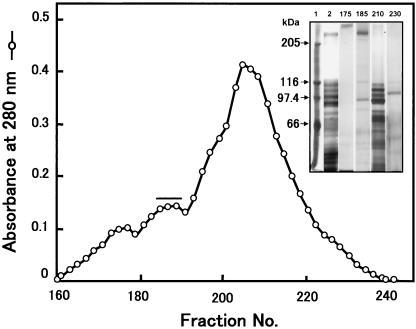
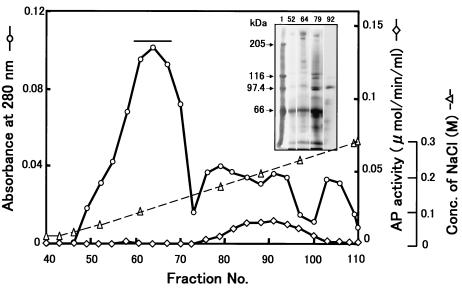
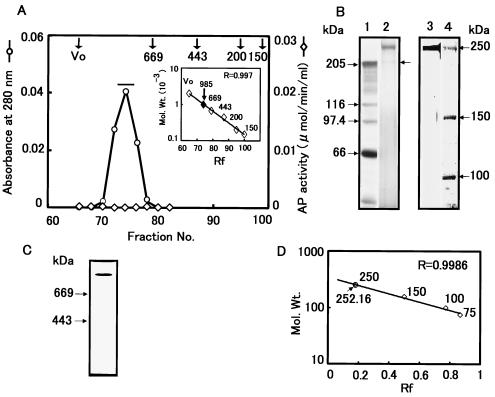
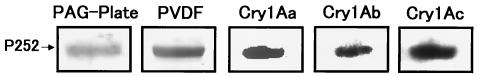
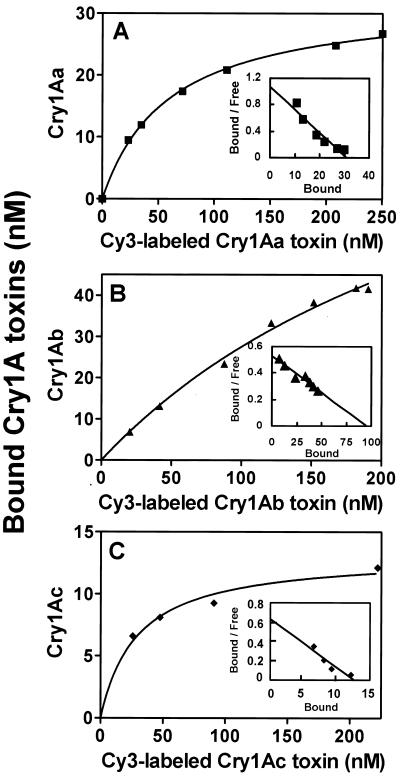
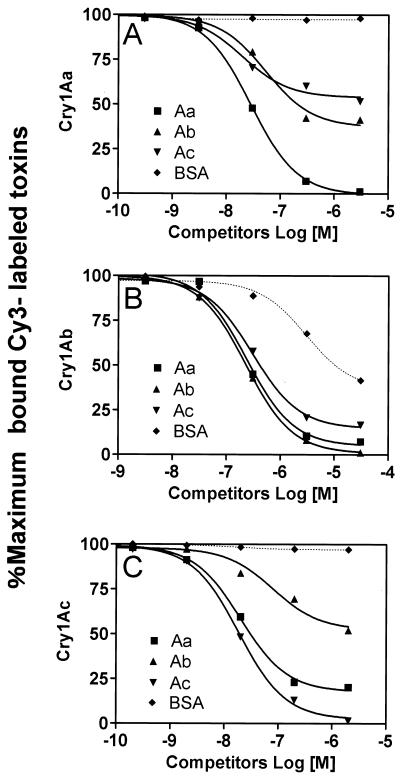
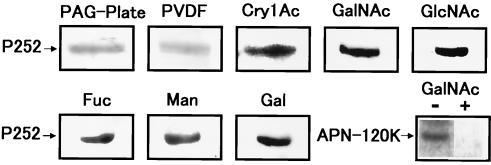
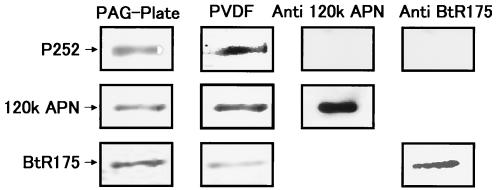
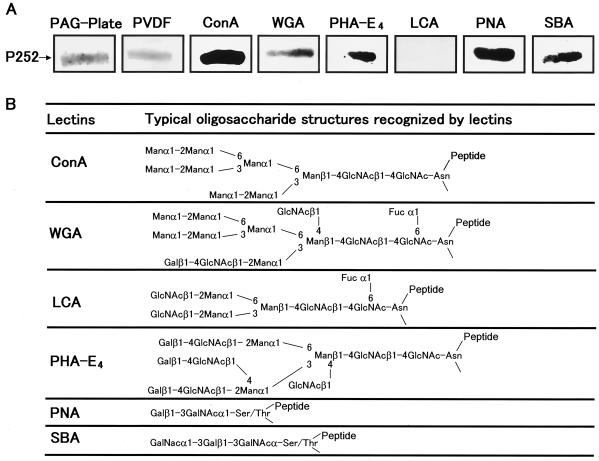
We describe the properties of a novel 252-kDa protein (P252) isolated from brush border membranes of Bombyx mori. P252 was found in a Triton X-100-soluble brush border membrane vesicle fraction, suggesting that it may be a component of the midgut epithelial cell membrane. P252 was purified to homogeneity, and the amino acid sequence of two internal peptides was determined, but neither of the peptides matched protein sequences in the available databases. The apparent molecular mass of the purified protein was estimated by denaturing gel electrophoresis to be 252 kDa, and it migrated as a single band on native gels. However, gel filtration chromatography indicated an apparent mass of 985 kDa, suggesting that P252 may exist as a homo-oligomer. The associations of P252 with Cry1Aa, Cry1Ab, and Cry1Ac were specific, and K(d) constants were determined to be 28.9, 178.5, and 20.0 nM, respectively. A heterologous competition assay was also done. P252 did not exhibit Leu-pNA hydrolysis activity, and binding to the Cry1A toxins was not inhibited by GalNAc. Binding assays of P252 with various lectins indicated the presence of three antennal N-linked high-mannose-type as well as O-linked mucin-type sugar side chains. While the function of P252 is not yet clear, we propose that it may function with Cry1A toxins during the insecticidal response and/or Cry toxin resistance mechanism.
Figures
References
-
- Baumann, N. A., and A. K. Menon. 2002. Lipid modification of proteins, p. 37-54. In D. E. Vance and J. E. Vance (ed.), Biochemistry of lipids, lipoproteins and membranes, 4th ed. Elsevier Sciences B. V., Amsterdam, The Netherlands.
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Cleveland, D. W., S. G. Fischer, M. L. Kirschner, and U. K. Laemmli. 1977. Peptide mapping by limited proteolysis in sodium dodecylsulphate and analysis by gel electrophoresis. J. Biol. Chem. 252:1102-1106. - PubMed
-
- Danielsen, E. M. 1994. Dimeric assembly of enterocyte brush border enzymes. Biochemistry 33:1599-1605. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
