

Natural stimulus statistics alter the receptive field structure of v1 neurons
- PMID: 15295035
- PMCID: PMC6729594
- DOI: 10.1523/JNEUROSCI.1422-04.2004
Natural stimulus statistics alter the receptive field structure of v1 neurons
Abstract
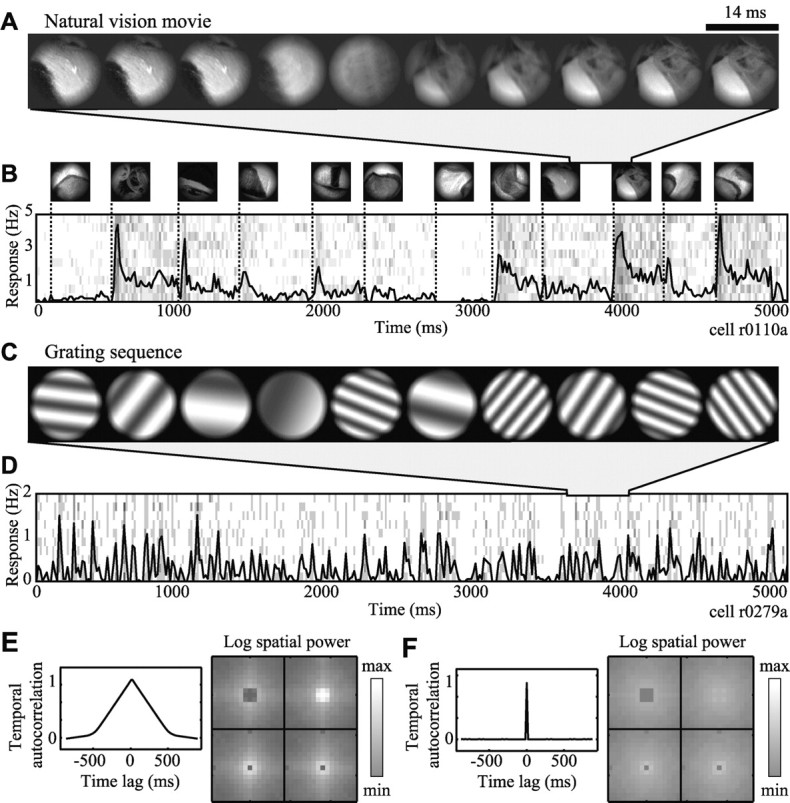
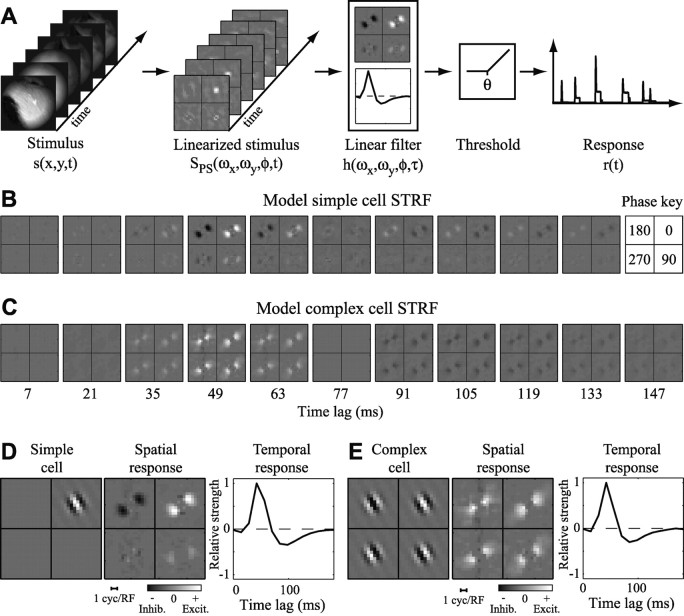
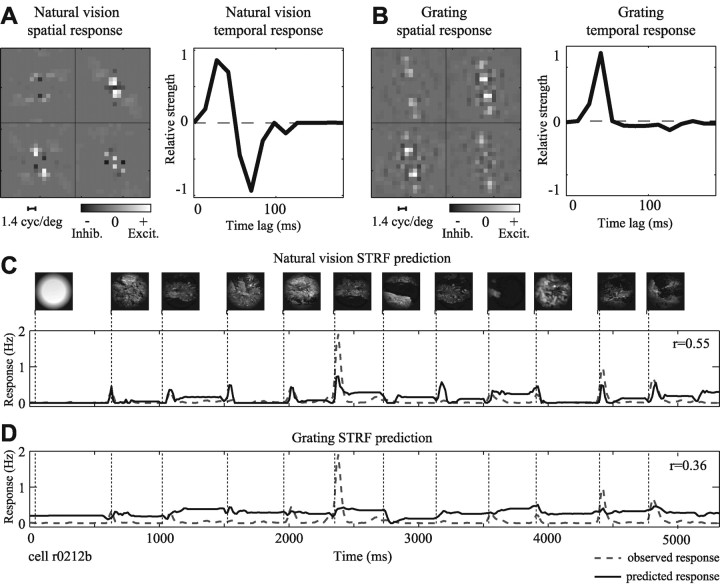
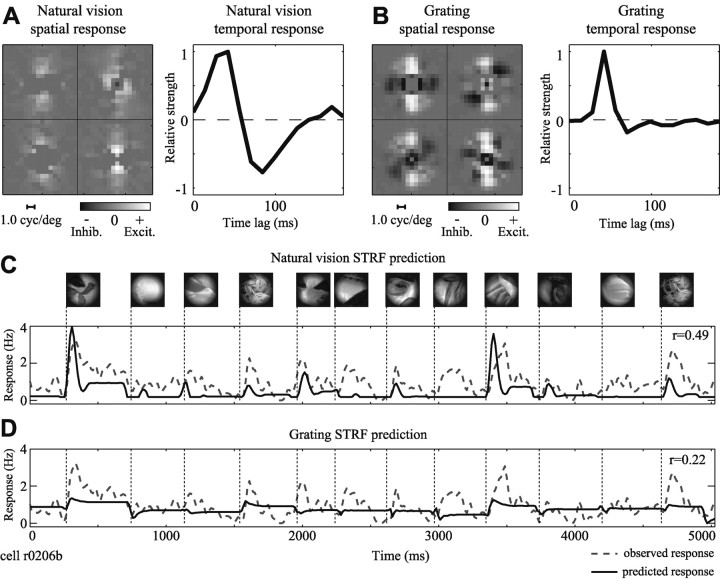
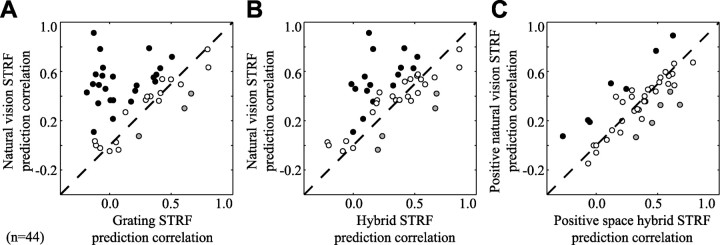
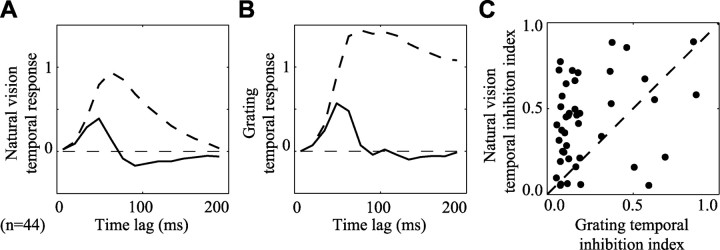
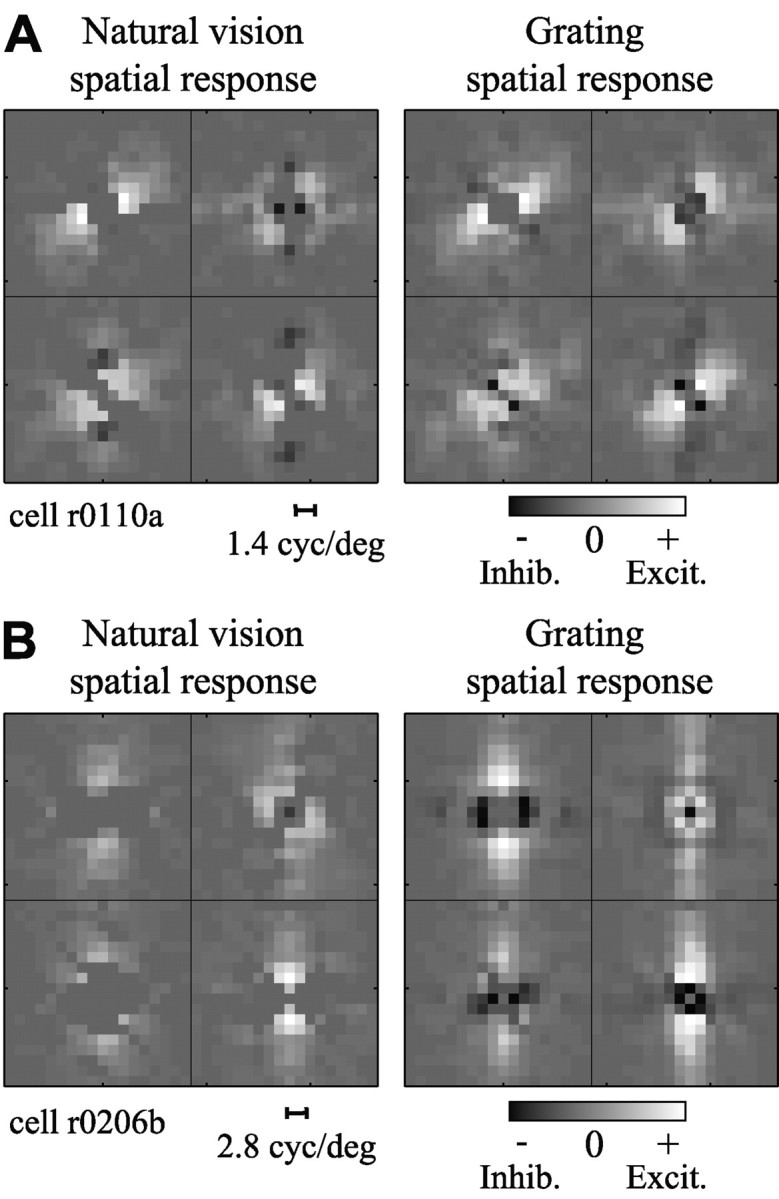
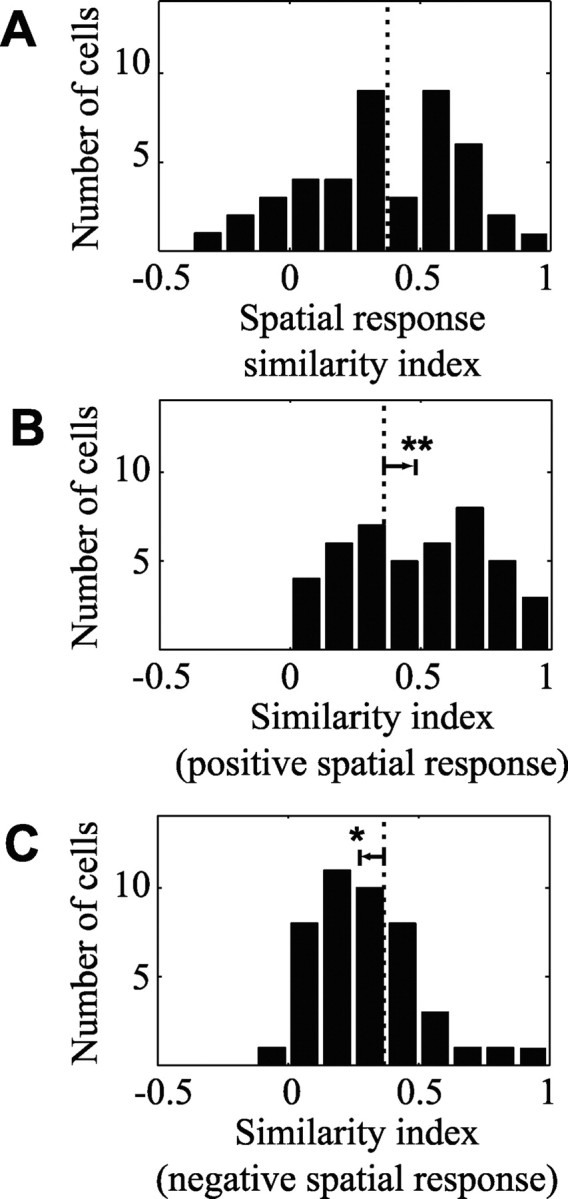
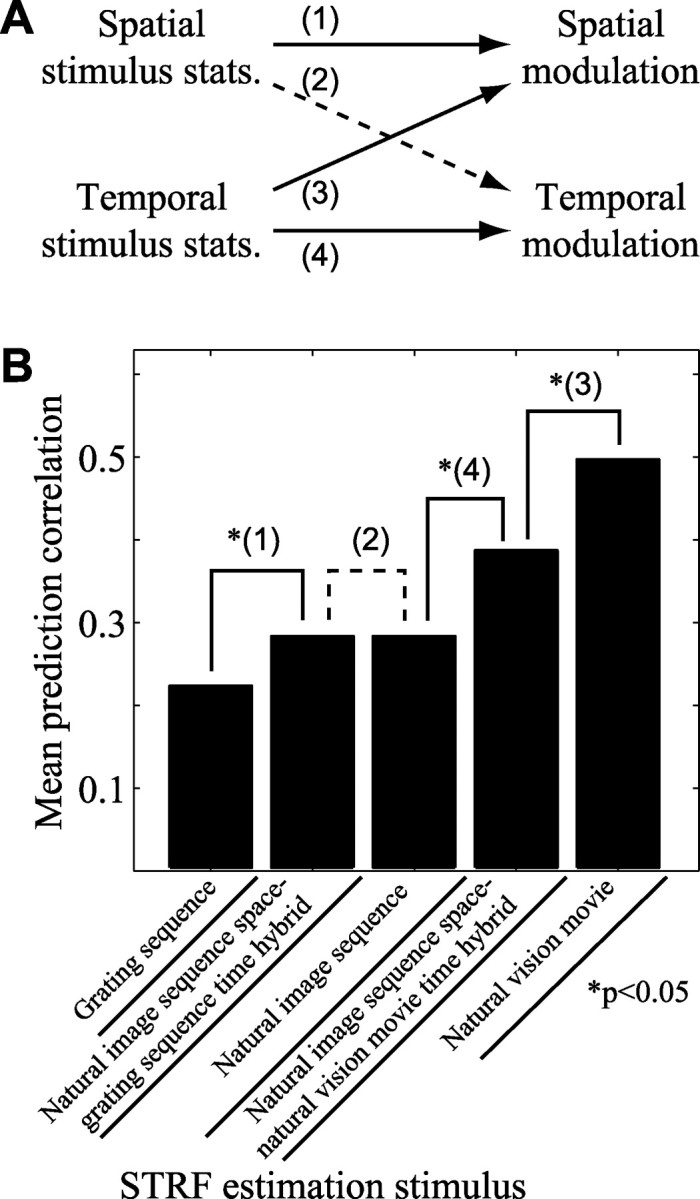
Studies of the primary visual cortex (V1) have produced models that account for neuronal responses to synthetic stimuli such as sinusoidal gratings. Little is known about how these models generalize to activity during natural vision. We recorded neural responses in area V1 of awake macaques to a stimulus with natural spatiotemporal statistics and to a dynamic grating sequence stimulus. We fit nonlinear receptive field models using each of these data sets and compared how well they predicted time-varying responses to a novel natural visual stimulus. On average, the model fit using the natural stimulus predicted natural visual responses more than twice as accurately as the model fit to the synthetic stimulus. The natural vision model produced better predictions in >75% of the neurons studied. This large difference in predictive power suggests that natural spatiotemporal stimulus statistics activate nonlinear response properties in a different manner than the grating stimulus. To characterize this modulation, we compared the temporal and spatial response properties of the model fits. During natural stimulation, temporal responses often showed a stronger late inhibitory component, indicating an effect of nonlinear temporal summation during natural vision. In addition, spatial tuning underwent complex shifts, primarily in the inhibitory, rather than excitatory, elements of the response profile. These differences in late and spatially tuned inhibition accounted fully for the difference in predictive power between the two models. Both the spatial and temporal statistics of the natural stimulus contributed to the modulatory effects.
Figures
References
-
- Adelson EH, Bergen JR (1985) Spatiotemporal energy models for the perception of motion. J Opt Soc Am 2: 284-299. - PubMed
-
- Aertsen AMHJ, Johannesma PIM (1981) A comparison of the spectro-temporal sensitivity of auditory neurons to tonal and natural stimuli. Biol Cybern 42: 145-156. - PubMed
-
- Albrecht DG, Geisler WS (1991) Motion selectivity and the contrast-response function of simple cells in the visual cortex. Vis Neurosci 7: 531-546. - PubMed
-
- Barlow HB (1961) Possible principles underlying the transformation of sensory messages. In: Sensory communication (Rosenbluth WA, ed), pp 217-234. Cambridge, MA: MIT.
-
- Bauman LA, Bonds AB (1991) Inhibitory refinement of spatial frequency selectivity in single cells of the cat striate cortex. Vision Res 31: 933-944. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources