

Fluorescence correlation spectroscopy relates rafts in model and native membranes
- PMID: 15298908
- PMCID: PMC1304444
- DOI: 10.1529/biophysj.104.040519
Fluorescence correlation spectroscopy relates rafts in model and native membranes
Abstract
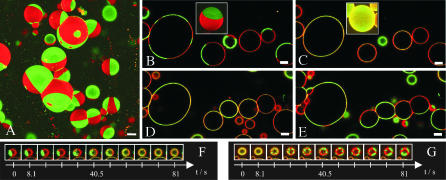
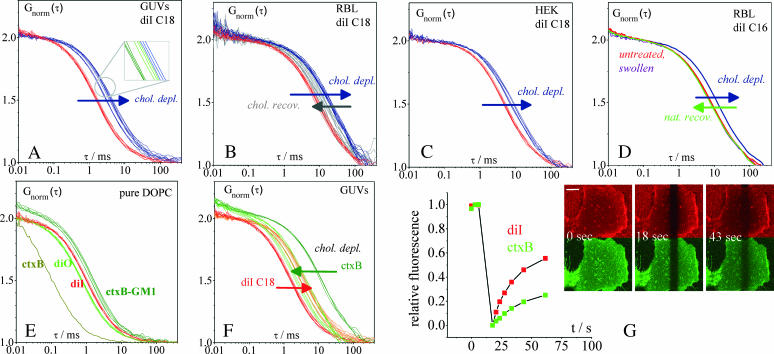
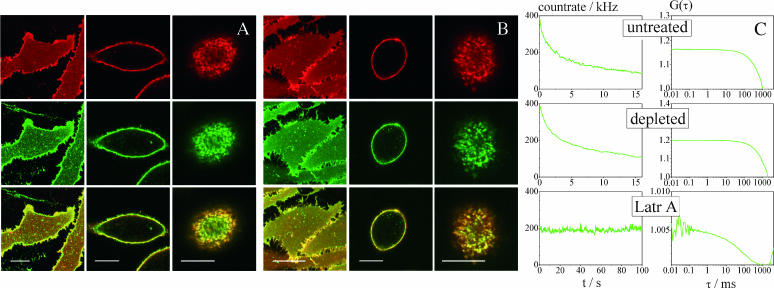
The lipid raft model has evoked a new perspective on membrane biology. Understanding the structure and dynamics of lipid domains could be a key to many crucial membrane-associated processes in cells. However, one shortcoming in the field is the lack of routinely applicable techniques to measure raft association without perturbation by detergents. We show that both in cell and in domain-exhibiting model membranes, fluorescence correlation spectroscopy (FCS) can easily distinguish a raft marker (cholera toxin B subunit bound to ganglioside (GM1) and a nonraft marker (dialkylcarbocyanine dye diI)) by their decidedly different diffusional mobilities. In contrast, these markers exhibit only slightly different mobilities in a homogeneous artificial membrane. Performing cholesterol depletion with methyl-beta-cyclodextrin, which disrupts raft organization, we find an analogous effect of reduced mobility for the nonraft marker in domain-exhibiting artificial membranes and in cell membranes. In contrast, cholesterol depletion has differential effects on the raft marker, cholera toxin B subunit-GM1, rendering it more mobile in artificial domain-exhibiting membranes but leaving it immobile in cell membranes, where cytoskeleton disruption is required to achieve higher mobility. Thus, fluorescence correlation spectroscopy promises to be a valuable tool to elucidate lipid raft associations in native cells and to gain deeper insight into the correspondence between model and natural membranes.
Figures
References
-
- Bacia, K., and P. Schwille. 2003. A dynamic view of cellular processes by in vivo fluorescence auto- and cross-correlation spectroscopy. Methods. 29:74–85. - PubMed
-
- Braccia, A., M. Villani, L. Immerdahl, L. L. Niels-Christiansen, B. T. Nystrom, G. H. Hansen, and E. M. Danielsen. 2003. Microvillar membrane microdomains exist at physiological temperature. Role of galectin-4 as lipid raft stabilizer revealed by “superrafts”. J. Biol. Chem. 278:15679–15684. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
