

doi: 10.1073/pnas.0402724101.
Epub 2004 Aug 6.
A mechanical basis for chromosome function
Affiliations
- PMID: 15299144
- PMCID: PMC515102
- DOI: 10.1073/pnas.0402724101
Item in Clipboard
A mechanical basis for chromosome function
Proc Natl Acad Sci U S A.
.
Abstract
We propose that chromosome function is governed by internal mechanical forces generated by programmed tendencies for expansion of the DNA/chromatin fiber against constraining features.
Figures
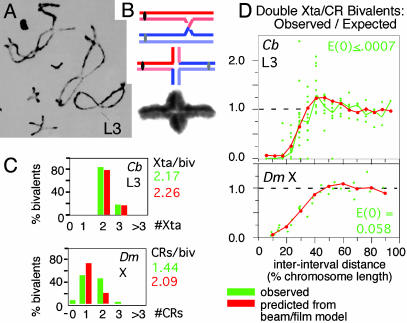
Chiasmata and crossovers. (A) Diplotene bivalents of Chorthippus brunneus with homologs linked by chiasmata (1). (B) Differential staining of sister chromatids shows that each chiasma is the site of a reciprocal exchange between one sister of each homolog (2). (C and D) Numbers and interference distributions of crossovers (D. melanogaster X chromosome; 3) and chiasmata (Chorthippus L3 bivalent, marked in A; ref. 1). Experimental values (green) and values predicted by the beam/film model (red; details in Fig. 10). [A and B are reproduced with permission from, respectively, ref. (Copyright 1984, Society for Experimental Biology) and ref. (Copyright 1978, Nature Publishing Group).]
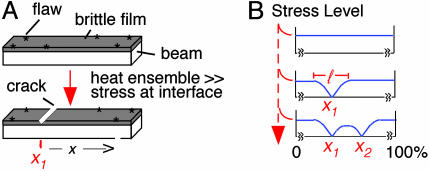
The beam/film system. See also text and Fig. 10.
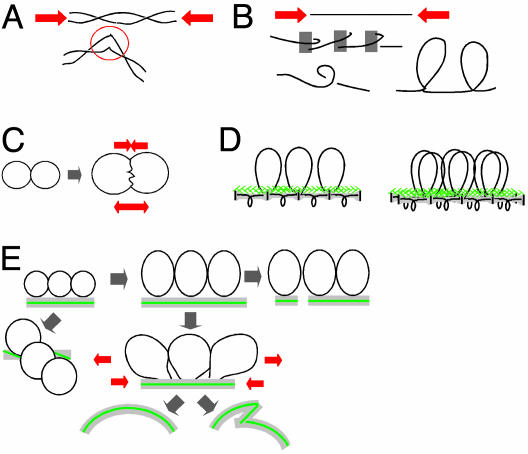
Stresses that could be generated by chromatin expansion against contraining features. (A and B) Compression stress along the DNA/chromatin fiber can destabilize DNA (A) or promote a more compact fiber configuration (B). (C) Interchromatin pushing produces opposing forces for separation and interface compression. (D) Linear/loop axis array (Left) and prophase configuration with cooriented sister arrays (Right). (E) Changes in axis status predicted from pushing forces between adjacent chromatin loops.
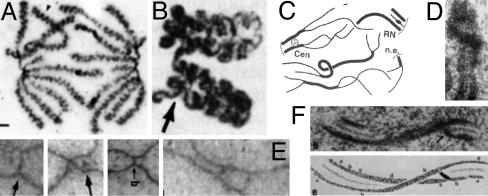
Observed changes in axis conformation. (A) Muntjac mitotic anaphase chromosomes with coiled axes (13). (B–F) Meiosis. (B) Coiled homologs of lily at diakinesis (14). (C) Axis fracturing and sister separation in late leptotene in Sordaria (15). (D) Twisting of a single leptotene homolog axis in Neotiella (16). (E)In Allium, coaligned homolog axes linked by bridges (Left) undergo apparent buckling plus ensuing nucleation of synaptonemal complex at late leptotene (1, 17). (F) Twisting synaptonemal complex (18). [Reproduced with permission from, respectively: (A) The Journal of Cell Biology, 1995, 131, pp. 7–17, by copyright permission of The Rockefeller University Press; (B) ref. (Copyright 1991, NRC Research Press); (C) ref. (Copyright 1999, Elsevier); (D) ref. by courtesy of the Carlsberg Laboratory (Copyright 1985, The Carlsberg Laboratory, Copenhagen); (E) ref. (Copyright 2004, Elsevier); and (F) ref. (Copyright 1978, NRC Research Press).]
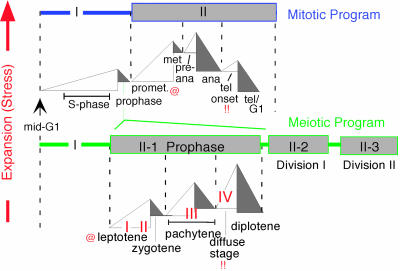
The mitotic and meiotic programs comprise related sequential cycles of global chromatin expansion and contraction, proposed to be cycles of expansional stress and stress relief. Four transitions of meiotic recombination (17) are indicated by I–IV. @ and!! symbolize analogies between prometaphase and telophase onset of the mitotic program and leptotene and diffuse stages of meiosis. @, appearance of silver-staining axis components along, and interaxis bridges between, sisters (mitosis, ref. 13) and homologs (meiosis, e.g., Fig. 4E) and chromosome destabilization in Sordaria spo76-1 (Fig. 14). (!!) Telophase onset and the diffuse stage both involve marked chromatin diffuseness and loss or destabilization of axial structures.
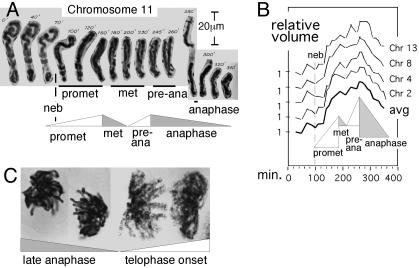
Chromatin expansion and contraction after prophase. (A and B) Individual Haemanthus chromosome volumes were determined from videomicrographs (e.g., A) (21). Shown are results for four chromosomes, normalized to 1 at t = 0 and their average. (C) At late anaphase, chromosomes are contracted, relaxed, and closely juxtaposed (Left); at telophase onset, chromosomes are expanded, distended, and separated (Right). [A and B are reproduced with permission from ref. (Copyright 1959, Blackwell Publishing, Ltd.).]
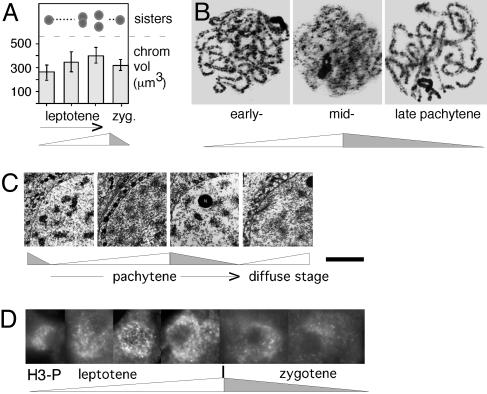
Chromatin expansion and contraction in meiotic prophase. (A) Chromatin volumes and sister chromatid cross-section morphologies seen by optical sectioning of maize leptotene and zygotene nuclei (25). (B) Pachytene chromosomes of Locusta migratoria (source, G.H.J). (C) Chromatin of human oocyte nuclei visualized by EM in ultrathin sections (26). At late zygotene/early pachytene, chromatin is highly condensed, leaving clear chromatin-free areas within the nucleus; at midpachytene, chromatin masses are less dense, and chromatin is more evenly distributed; compact chromatin returns at late pachytene; and less dense chromatin then returns at “the diffuse stage.” (D) Immunostaining of histone H3 Ser-10 phosphorylation at successive stages of meiotic leptotene/zygotene in Sordaria (source, D.Z.; Fig. 11). [A and C are reprinted with permission from, respectively, ref. (Copyright 1994, Elsevier) and ref. by courtesy of the Carlsberg Laboratory (Copyright 1970, The Carlsberg Laboratory, Copenhagen).]
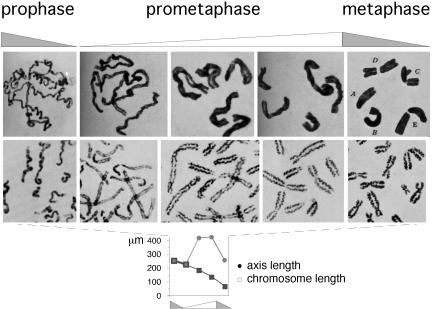
Chromosomes at prometaphase/metaphase. (Top) Mitotic chromosomes of T. grandiflorum (acetocarmine preparations; figures 19–24 of ref. 29). (Middle and Bottom) Human chromosomes stained to reveal axes (30) and end-to-end and axis lengths for corresponding total chromosome complements (details in Supporting Text, which is published as supporting information on the PNAS web site). [Reproduced with permission from, respectively: (Top) ref. (Copyright 1941, NRC Research Press); (Middle and Bottom) ref. (Copyright 1968, Springer-Verlag GmbH & Co.).]
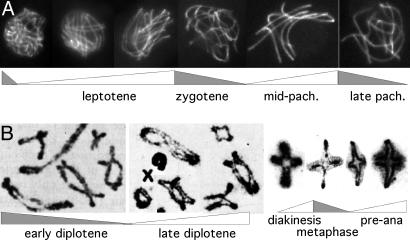
Modulations of meiotic prophase chromosome conformation in correlation with chromatin expansion/contraction cycles. (A) Sordaria chromosome axes stained with Spo76-GFP are stiffer or floppier in periods of expansion or contraction, respectively (source, D.Z.). (B) Whole nuclear complements of locust chromosomes at early (Left) and late (Right) diplotene exhibit conjoined and separated sister chromatids (34). (C) Individual grasshopper chromosomes showing stained chromatid axes (35) alternating between conjoined and separated configurations during periods ofcontraction and expansion, respectively. Additional examples are in Figs. 11 A and B and 13. [B is reproduced from ref. ; C is reproduced with permission from ref. (Copyright 1990, NRC Research Press).]
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
