

Major genetic marker of nidoviruses encodes a replicative endoribonuclease
- PMID: 15304651
- PMCID: PMC514660
- DOI: 10.1073/pnas.0403127101
Major genetic marker of nidoviruses encodes a replicative endoribonuclease
Abstract
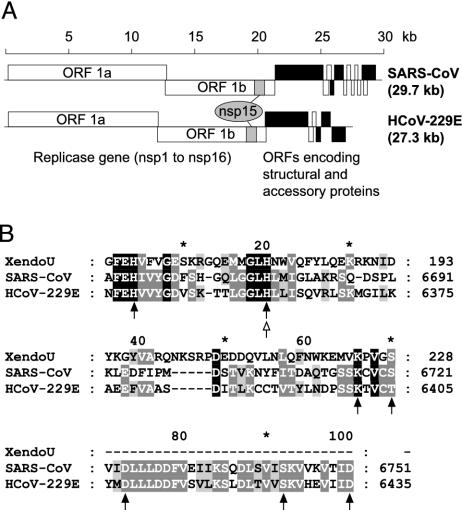
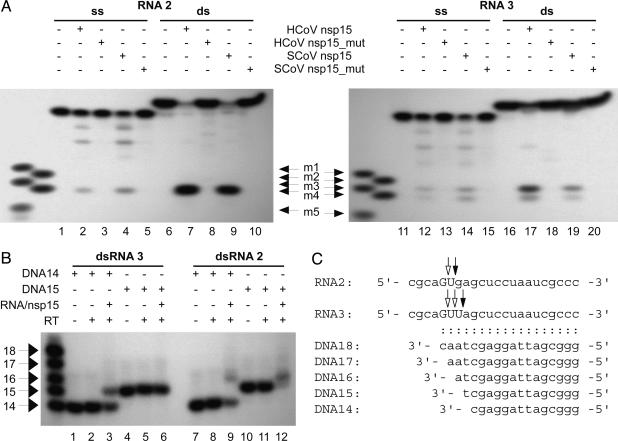
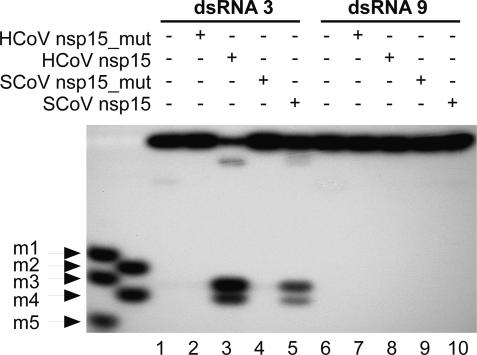
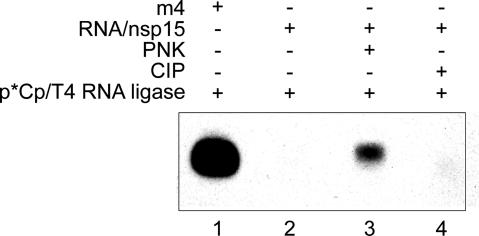
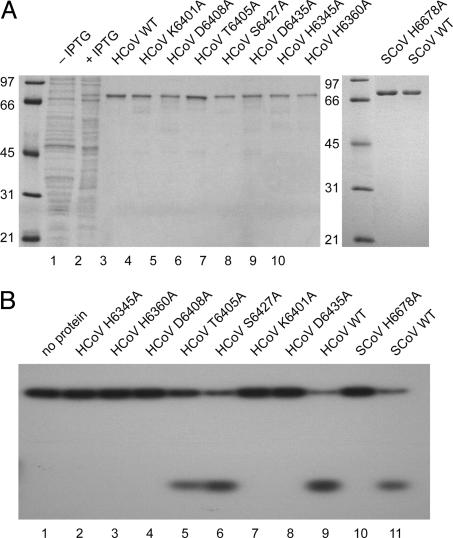
Coronaviruses are important pathogens that cause acute respiratory diseases in humans. Replication of the approximately 30-kb positive-strand RNA genome of coronaviruses and discontinuous synthesis of an extensive set of subgenome-length RNAs (transcription) are mediated by the replicase-transcriptase, a barely characterized protein complex that comprises several cellular proteins and up to 16 viral subunits. The coronavirus replicase-transcriptase was recently predicted to contain RNA-processing enzymes that are extremely rare or absent in other RNA viruses. Here, we established and characterized the activity of one of these enzymes, replicative nidoviral uridylate-specific endoribonuclease (NendoU). It is considered a major genetic marker that discriminates nidoviruses (Coronaviridae, Arteriviridae, and Roniviridae) from all other RNA virus families. Bacterially expressed forms of NendoU of severe acute respiratory syndrome coronavirus and human coronavirus 229E were revealed to cleave single-stranded and double-stranded RNA in a Mn(2+)-dependent manner. Single-stranded RNA was cleaved less specifically and effectively, suggesting that double-stranded RNA is the biologically relevant NendoU substrate. Double-stranded RNA substrates were cleaved upstream and downstream of uridylates at GUU or GU sequences to produce molecules with 2'-3' cyclic phosphate ends. 2'-O-ribose-methylated RNA substrates proved to be resistant to cleavage by NendoU, indicating a functional link with the 2'-O-ribose methyltransferase located adjacent to NendoU in the coronavirus replicative polyprotein. A mutagenesis study verified potential active-site residues and allowed us to inactivate NendoU in the full-length human coronavirus 229E clone. Substitution of D6408 by Ala was shown to abolish viral RNA synthesis, demonstrating that NendoU has critical functions in viral replication and transcription.
Figures

References
-
- Myint, S. H. (1995) in The Coronaviridae, ed. Siddell, S. G. (Plenum, New York), pp. 389-401.
-
- Peiris, J. S., Yuen, K. Y., Osterhaus, A. D. & Stöhr, K. (2003) N. Engl. J. Med. 349, 2431-2441. - PubMed
-
- Thiel, V., Ivanov, K. A., Putics, A., Hertzig, T., Schelle, B., Bayer, S., Weissbrich, B., Snijder, E. J., Rabenau, H., Doerr, H. W., Gorbalenya, A. E. & Ziebuhr, J. (2003) J. Gen. Virol. 84, 2305-2315. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
