

Genome-wide searching for pseudouridylation guide snoRNAs: analysis of the Saccharomyces cerevisiae genome
- PMID: 15306656
- PMCID: PMC514388
- DOI: 10.1093/nar/gkh768
Genome-wide searching for pseudouridylation guide snoRNAs: analysis of the Saccharomyces cerevisiae genome
Abstract
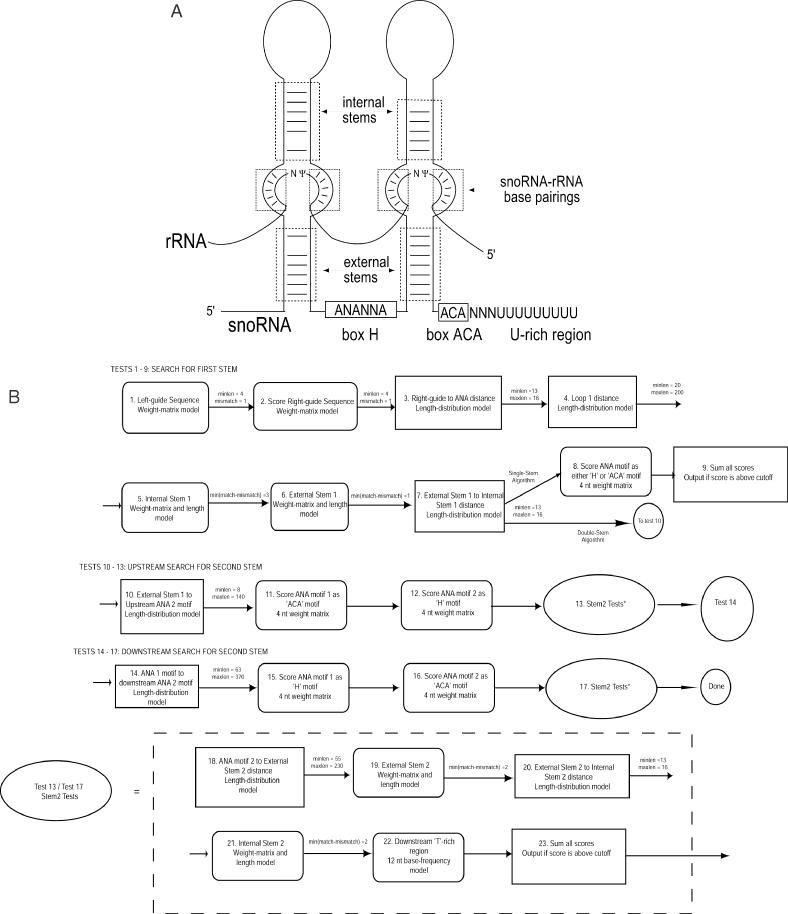
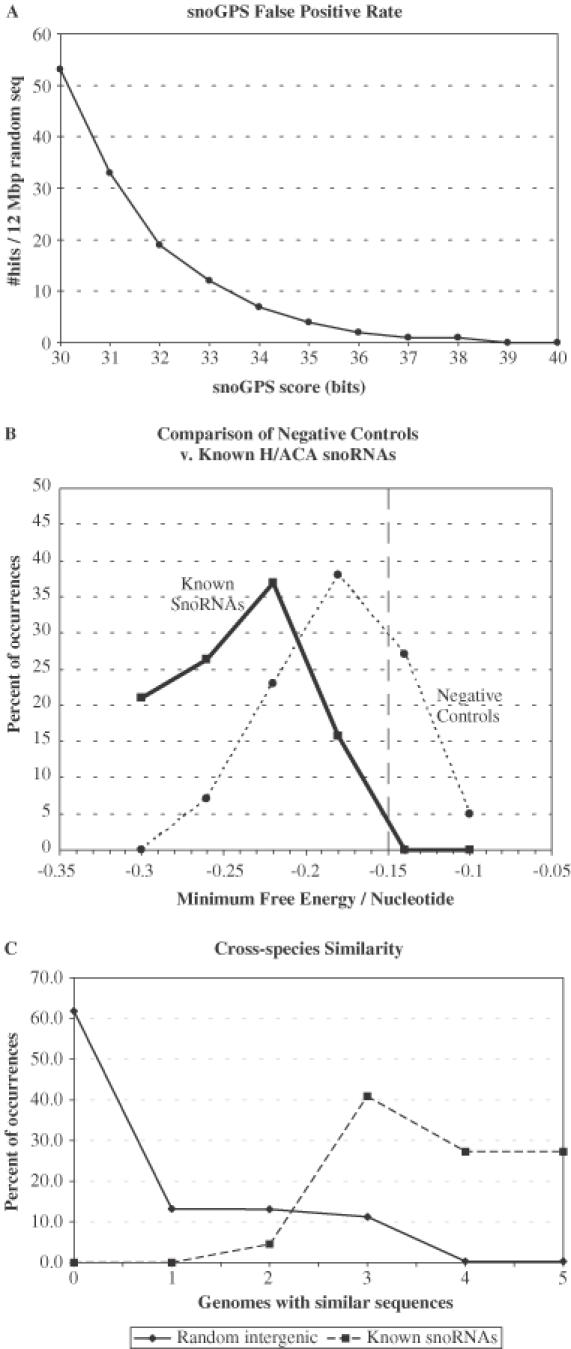
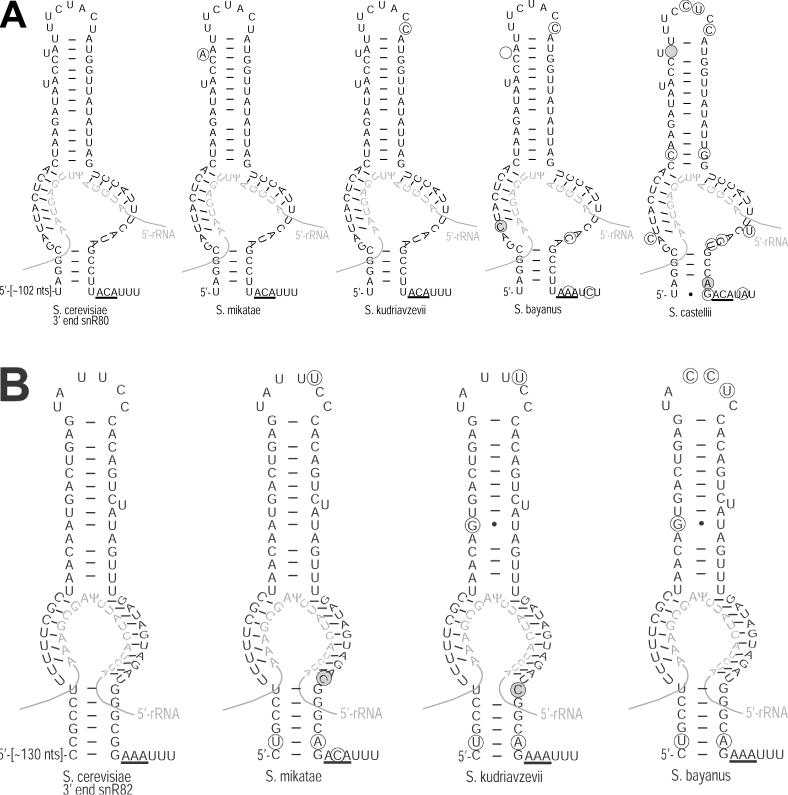
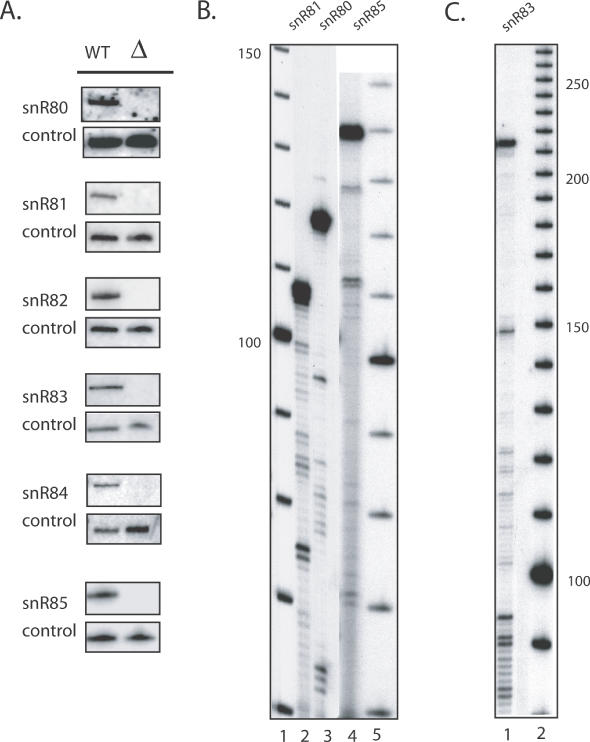
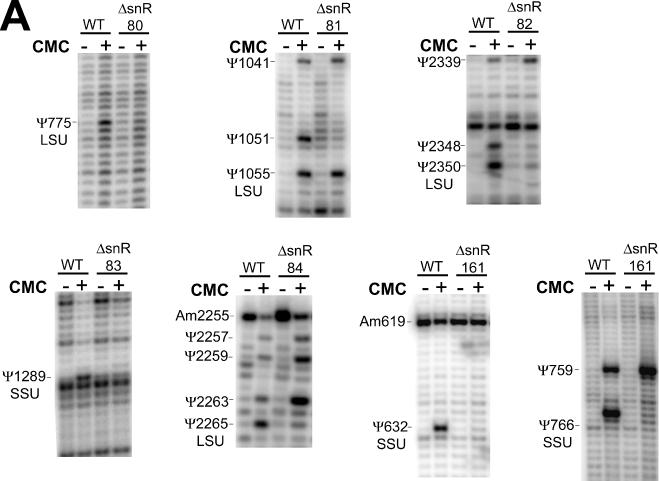
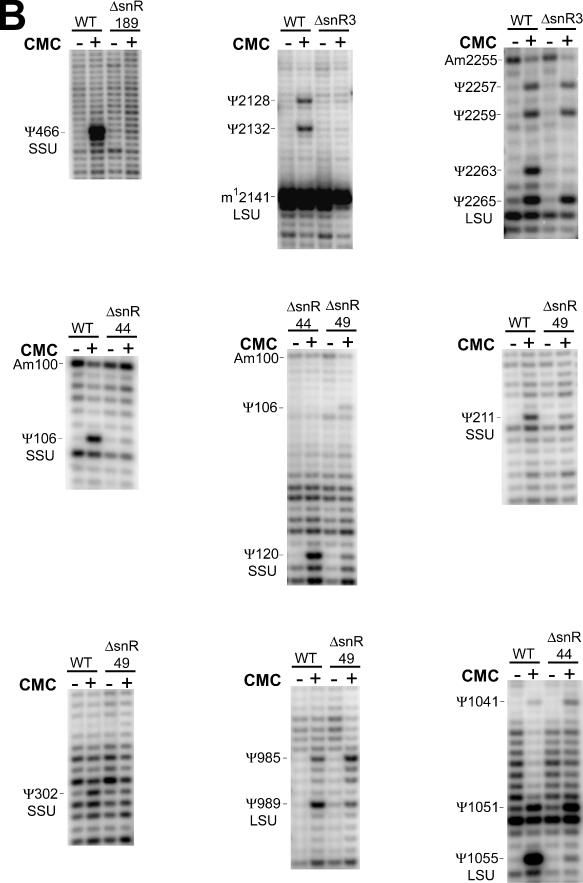
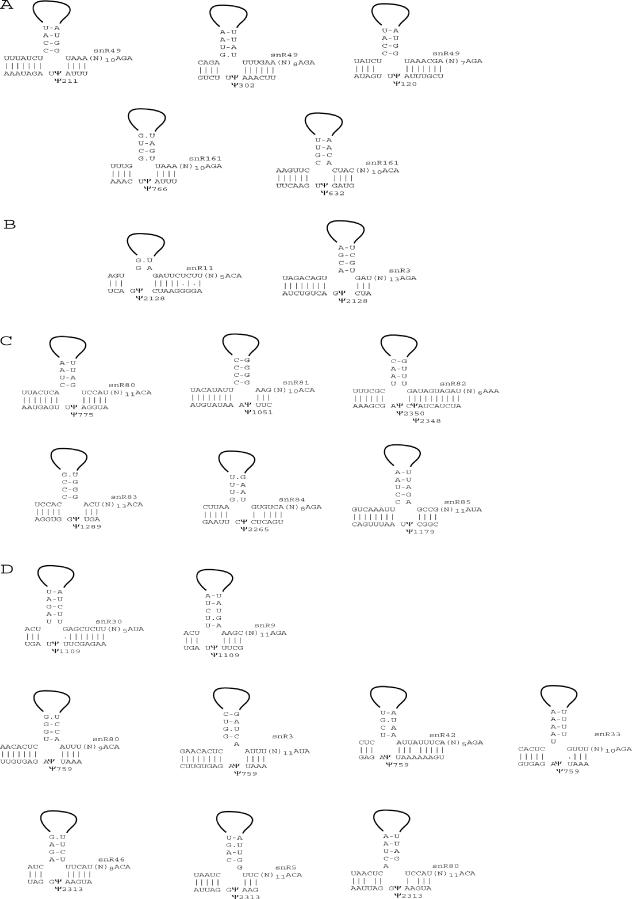
One of the largest families of small RNAs in eukaryotes is the H/ACA small nucleolar RNAs (snoRNAs), most of which guide RNA pseudouridine formation. So far, an effective computational method specifically for identifying H/ACA snoRNA gene sequences has not been established. We have developed snoGPS, a program for computationally screening genomic sequences for H/ACA guide snoRNAs. The program implements a deterministic screening algorithm combined with a probabilistic model to score gene candidates. We report here the results of testing snoGPS on the budding yeast Saccharomyces cerevisiae. Six candidate snoRNAs were verified as novel RNA transcripts, and five of these were verified as guides for pseudouridine formation at specific sites in ribosomal RNA. We also predicted 14 new base-pairings between snoRNAs and known pseudouridine sites in S.cerevisiae rRNA, 12 of which were verified by gene disruption and loss of the cognate pseudouridine site. Our findings include the first prediction and verification of snoRNAs that guide pseudouridine modification at more than two sites. With this work, 41 of the 44 known pseudouridine modifications in S.cerevisiae rRNA have been linked with a verified snoRNA, providing the most complete accounting of the H/ACA snoRNAs that guide pseudouridylation in any species.
Figures
Similar articles
-
The complete set of H/ACA snoRNAs that guide rRNA pseudouridylations in Saccharomyces cerevisiae.RNA. 2005 Jun;11(6):928-38. doi: 10.1261/rna.2100905. RNA. 2005. PMID: 15923376 Free PMC article.
-
A computational screen for mammalian pseudouridylation guide H/ACA RNAs.RNA. 2006 Jan;12(1):15-25. doi: 10.1261/rna.2210406. RNA. 2006. PMID: 16373490 Free PMC article.
-
A snoRNA that guides the two most conserved pseudouridine modifications within rRNA confers a growth advantage in yeast.RNA. 2003 Jul;9(7):771-9. doi: 10.1261/rna.5240503. RNA. 2003. PMID: 12810910 Free PMC article.
-
Small RNAs with big implications: new insights into H/ACA snoRNA function and their role in human disease.Wiley Interdiscip Rev RNA. 2015 Mar-Apr;6(2):173-89. doi: 10.1002/wrna.1266. Epub 2014 Oct 31. Wiley Interdiscip Rev RNA. 2015. PMID: 25363811 Free PMC article. Review.
-
The expanding snoRNA world.Biochimie. 2002 Aug;84(8):775-90. doi: 10.1016/s0300-9084(02)01402-5. Biochimie. 2002. PMID: 12457565 Review.
Cited by
-
Comparative genomics of eukaryotic small nucleolar RNAs reveals deep evolutionary ancestry amidst ongoing intragenomic mobility.BMC Evol Biol. 2012 Sep 15;12:183. doi: 10.1186/1471-2148-12-183. BMC Evol Biol. 2012. PMID: 22978381 Free PMC article.
-
Enhanced leavening ability of baker's yeast by overexpression of SNR84 with PGM2 deletion.J Ind Microbiol Biotechnol. 2015 Jun;42(6):939-48. doi: 10.1007/s10295-015-1618-5. Epub 2015 Apr 16. J Ind Microbiol Biotechnol. 2015. PMID: 25877163
-
A comparative genome-wide study of ncRNAs in trypanosomatids.BMC Genomics. 2010 Nov 4;11:615. doi: 10.1186/1471-2164-11-615. BMC Genomics. 2010. PMID: 21050447 Free PMC article.
-
Discovery of Pyrobaculum small RNA families with atypical pseudouridine guide RNA features.RNA. 2012 Mar;18(3):402-11. doi: 10.1261/rna.031385.111. Epub 2012 Jan 26. RNA. 2012. PMID: 22282340 Free PMC article.
-
5'-End sequencing in Saccharomyces cerevisiae offers new insights into 5'-ends of tRNAHis and snoRNAs.FEBS Lett. 2019 May;593(9):971-981. doi: 10.1002/1873-3468.13364. Epub 2019 Apr 4. FEBS Lett. 2019. PMID: 30908619 Free PMC article.
References
-
- Bachellerie J.P., Cavaille,J. and Huttenhofer,A. (2002) The expanding snoRNA world. Biochimie, 84, 775–790. - PubMed
-
- Bertrand E. and Fournier,M.J. (2004) The snoRNPs and related machines: ancient devices that mediate maturation of rRNA and other RNAs. In Olson,M. (ed.), The Nucleolus. Landes Bioscience, Georgetown, TX, pp. 225–261.
-
- Kiss T. (2002) Small nucleolar RNAs: an abundant group of noncoding RNAs with diverse cellular functions. Cell, 109, 145–148. - PubMed
-
- Lowe T.M. and Eddy,S.R. (1999) A computational screen for methylation guide snoRNAs in yeast. Science, 283, 1168–1171. - PubMed
-
- Omer A.D., Lowe,T.M., Russell,A.G., Ebhardt,H., Eddy,S.R. and Dennis,P.P. (2000) Homologs of small nucleolar RNAs in Archaea. Science, 288, 517–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
