

Structural basis for proteolysis-dependent activation of the poliovirus RNA-dependent RNA polymerase
- PMID: 15306852
- PMCID: PMC516629
- DOI: 10.1038/sj.emboj.7600357
Structural basis for proteolysis-dependent activation of the poliovirus RNA-dependent RNA polymerase
Abstract
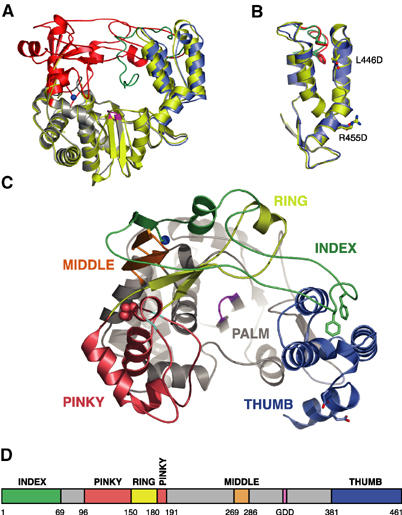
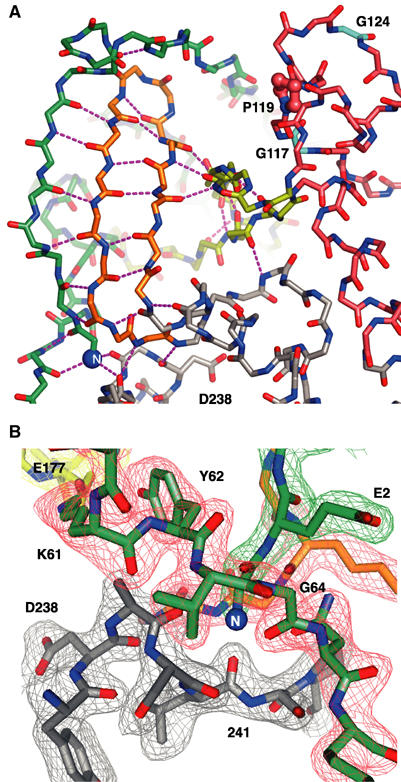
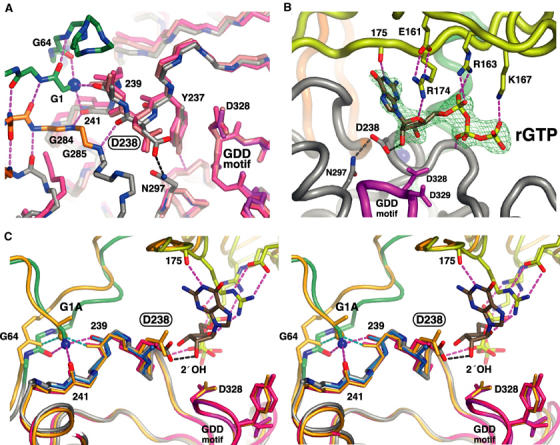
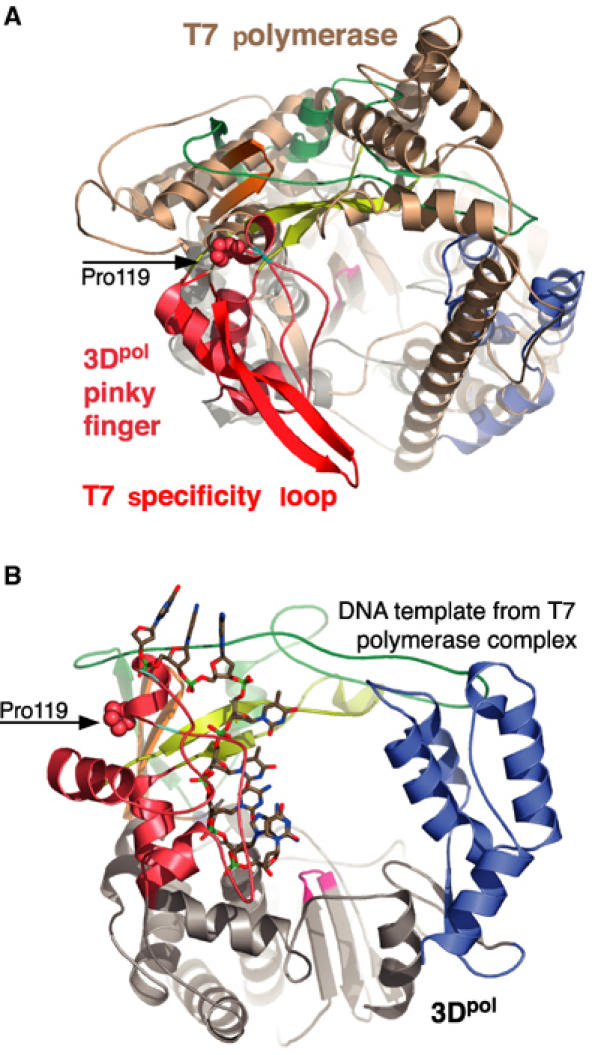
The active RNA-dependent RNA polymerase of poliovirus, 3Dpol, is generated by cleavage of the 3CDpro precursor protein, a protease that has no polymerase activity despite containing the entire polymerase domain. By intentionally disrupting a known and persistent crystal packing interaction, we have crystallized the poliovirus polymerase in a new space group and solved the complete structure of the protein at 2.0 A resolution. It shows that the N-terminus of fully processed 3Dpol is buried in a surface pocket where it makes hydrogen bonds that act to position Asp238 in the active site. Asp238 is an essential residue that selects for the 2' OH group of substrate rNTPs, as shown by a 2.35 A structure of a 3Dpol-GTP complex. Mutational, biochemical, and structural data further demonstrate that 3Dpol activity is exquisitely sensitive to mutations at the N-terminus. This sensitivity is the result of allosteric effects where the structure around the buried N-terminus directly affects the positioning of Asp238 in the active site.
Figures
References
-
- Ago H, Adachi T, Yoshida A, Yamamoto M, Habuka N, Yatsunami K, Miyano M (1999) Crystal structure of the RNA-dependent RNA polymerase of hepatitis C virus. Struct Fold Des 7: 1417–1426 - PubMed
-
- Andreotti AH (2003) Native state proline isomerization: an intrinsic molecular switch. Biochemistry 42: 9515–9524 - PubMed
-
- Arnold JJ, Cameron CE (2000) Poliovirus RNA-dependent RNA polymerase 3D(pol). Assembly of stable, elongation-competent complexes by using a symmetrical primer–template substrate (sym/sub). J Biol Chem 275: 5329–5336 - PubMed
-
- Brandts JF, Halvorson HR, Brennan M (1975) Consideration of the possibility that the slow step in protein denaturation reactions is due to cis–trans isomerism of proline residues. Biochemistry 14: 4953–4963 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
