

Analysis of human immunodeficiency virus type 1 transcriptional elongation in resting CD4+ T cells in vivo
- PMID: 15308706
- PMCID: PMC506937
- DOI: 10.1128/JVI.78.17.9105-9114.2004
Analysis of human immunodeficiency virus type 1 transcriptional elongation in resting CD4+ T cells in vivo
Abstract
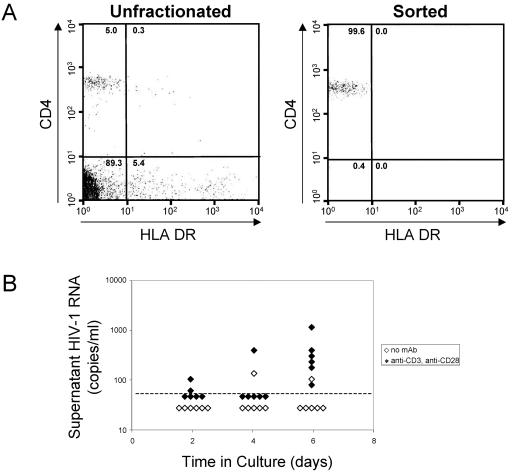
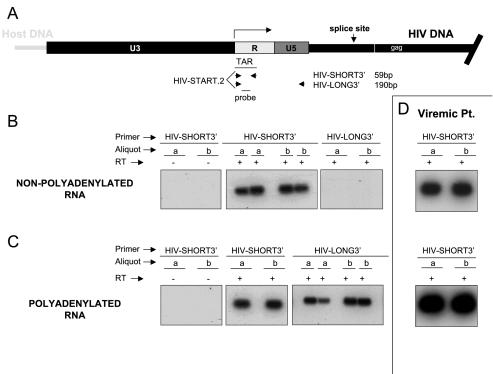
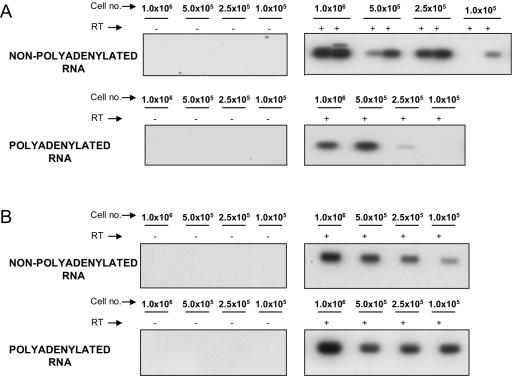
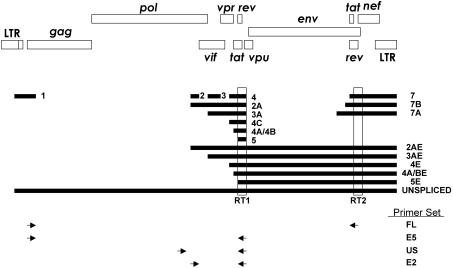
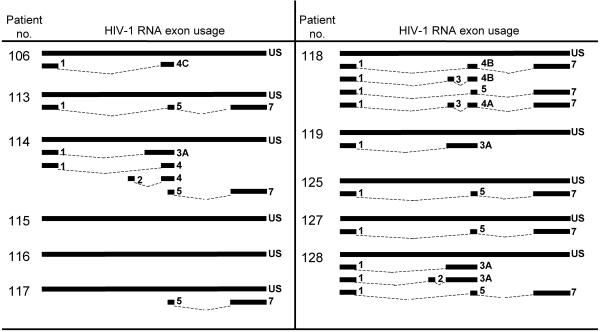
A stable latent reservoir for human immunodeficiency virus type 1 (HIV-1) in resting memory CD4+ T cells presents a barrier to eradication of the infection even in patients on highly active antiretroviral therapy. Potential mechanisms for latency include inaccessibility of the integrated viral genome, absence of key host transcription factors, premature termination of HIV-1 RNAs, and abnormal splicing patterns. To differentiate among these mechanisms, we isolated extremely pure populations of resting CD4+ T cells from patients on highly active antiretroviral therapy. These cells did not produce virus but retained the capacity to do so if appropriately stimulated. Products of HIV-1 transcription were examined in purified resting CD4+ T cells. Although short, prematurely terminated HIV-1 transcripts have been suggested as a marker for latently infected cells, the production of short transcripts had not been previously demonstrated in purified populations of resting CD4+ T cells. By separating RNA into polyadenylated and nonpolyadenylated fractions, we showed that resting CD4+ T cells from patients on highly active antiretroviral therapy produce abortive transcripts that lack a poly(A) tail and that terminate prior to nucleotide 181. Short transcripts dominated the pool of total HIV-1 transcripts in resting CD4+ T cells. Processive, polyadenylated HIV-1 mRNAs were also present at a low level. Both unspliced and multiply spliced forms were found. Taken together, these results show that the nonproductive nature of the infection in resting CD4+ T cells from patients on highly active antiretroviral therapy is not due to absolute blocks at the level of either transcriptional initiation or elongation but rather relative inefficiencies at multiple steps.
Figures
References
-
- Adams, M., L. Sharmeen, J. Kimpton, J. M. Romeo, J. V. Garcia, B. M. Peterlin, M. Groudine, and M. Emerman. 1994. Cellular latency in human immunodeficiency virus-infected individuals with high CD4 levels can be detected by the presence of promoter-proximal transcripts. Proc. Natl. Acad. Sci. USA 91:3862-3866. - PMC - PubMed
-
- Adams, M., C. Wong, D. Wang, and J. Romeo. 1999. Limitation of Tat-associated transcriptional processivity in HIV-infected PBMC. Virology 257:397-405. - PubMed
-
- Alcami, J., D. L. Lain, L. Folgueira, M. A. Pedraza, J. M. Jacque, F. Bachelerie, A. R. Noriega, R. T. Hay, D. Harrich, and R. B. Gaynor. 1995. Absolute dependence on kappa B responsive elements for initiation and Tat-mediated amplification of HIV transcription in blood CD4 T lymphocytes. EMBO J. 14:1552-1560. - PMC - PubMed
-
- Bagnarelli, P., A. Valenza, S. Menzo, R. Sampaolesi, P. E. Varaldo, L. Butini, M. Montroni, C. F. Perno, S. Aquaro, D. Mathez, J. Leibowitch, C. Balotta, and M. Clementi. 1996. Dynamics and modulation of human immunodeficiency virus type 1 transcripts in vitro and in vivo. J. Virol. 70:7603-7613. - PMC - PubMed
-
- Barboric, M., R. M. Nissen, S. Kanazawa, N. Jabrane-Ferrat, and B. M. Peterlin. 2001. NF-kappaB binds P-TEFb to stimulate transcriptional elongation by RNA polymerase II. Mol. Cell 8:327-337. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
