

Identification of cis sequences required for lytic DNA replication and packaging of murine gammaherpesvirus 68
- PMID: 15308708
- PMCID: PMC506910
- DOI: 10.1128/JVI.78.17.9123-9131.2004
Identification of cis sequences required for lytic DNA replication and packaging of murine gammaherpesvirus 68
Abstract
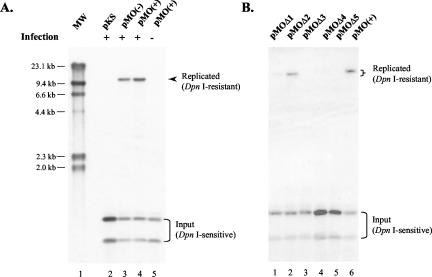
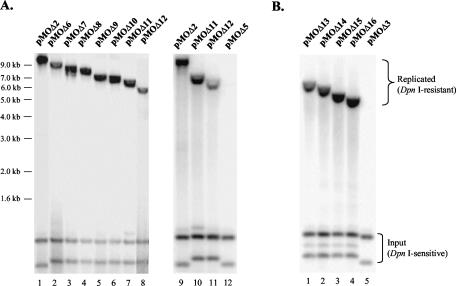
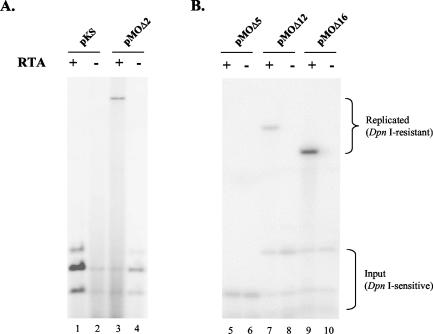
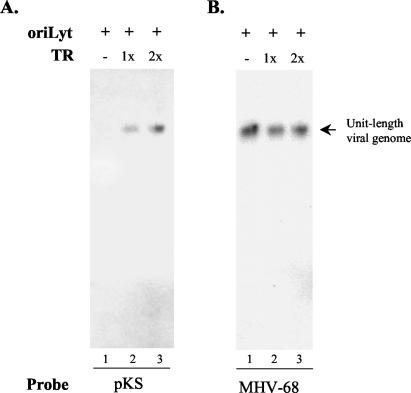
Human gammaherpesviruses are associated with lymphomas and other malignancies. Murine gammaherpesvirus 68 (MHV-68) infection of mice has emerged as a model for understanding gammaherpesvirus pathogenesis in vivo. In contrast to human gammaherpesviruses, MHV-68 replicates in permissive cell lines in a robust manner, presenting an efficient model to study the basic mechanisms for DNA replication and recombination processes. In addition, MHV-68 also infects a broad range of cells of different tissue types and from different host species, and the viral genome persists as an episome in infected cells. These features make MHV-68 an attractive system on which to build gene delivery vectors. We have therefore undertaken a study to identify the cis elements required for MHV-68 genome replication and packaging. Here we report that an 8.4-kb MHV-68 genomic fragment between ORF66 and ORF73 conferred on the plasmid the ability to replicate; replication required the presence of either de novo viral infection or viral reactivation from latency. We further mapped the origin of lytic replication (oriLyt) to a 1.25-kb region. Moreover, we demonstrated that the terminal repeat of the viral genome is sufficient for packaging of the replicated oriLyt plasmid into mature viral particles. Functional identification of the MHV-68 oriLyt and packaging signal has laid a foundation for investigating the mechanisms controlling gammaherpesvirus DNA replication during the viral lytic phase and will also serve as a base on which to design gene delivery vectors.
Figures

Similar articles
-
CCAAT/enhancer binding proteins play a role in oriLyt-dependent genome replication during MHV-68 de novo infection.Protein Cell. 2011 Jun;2(6):463-9. doi: 10.1007/s13238-011-1060-z. Epub 2011 Jul 12. Protein Cell. 2011. PMID: 21748596 Free PMC article.
-
Identification and functional characterization of the left origin of lytic replication of murine gammaherpesvirus 68.Virology. 2009 May 10;387(2):285-95. doi: 10.1016/j.virol.2009.02.029. Epub 2009 Mar 12. Virology. 2009. PMID: 19285330 Free PMC article.
-
Murine Gammaherpesvirus 68 ORF48 Is an RTA-Responsive Gene Product and Functions in both Viral Lytic Replication and Latency during In Vivo Infection.J Virol. 2015 Jun;89(11):5788-800. doi: 10.1128/JVI.00406-15. Epub 2015 Mar 11. J Virol. 2015. PMID: 25762743 Free PMC article.
-
Is murine gammaherpesvirus-68 (MHV-68) a suitable immunotoxicological model for examining immunomodulatory drug-associated viral recrudescence?J Immunotoxicol. 2015 Jan-Mar;12(1):1-15. doi: 10.3109/1547691X.2014.882996. Epub 2014 Feb 10. J Immunotoxicol. 2015. PMID: 24512328 Review.
-
Plasmid Partitioning by Human Tumor Viruses.J Virol. 2018 Apr 13;92(9):e02170-17. doi: 10.1128/JVI.02170-17. Print 2018 May 1. J Virol. 2018. PMID: 29467315 Free PMC article. Review.
Cited by
-
Replication and transcription activator (RTA) of murine gammaherpesvirus 68 binds to an RTA-responsive element and activates the expression of ORF18.J Virol. 2011 Nov;85(21):11338-50. doi: 10.1128/JVI.00561-11. Epub 2011 Aug 17. J Virol. 2011. PMID: 21849436 Free PMC article.
-
Dynamic association of gammaherpesvirus DNA with core histone during de novo lytic infection of primary cells.Virology. 2011 Dec 20;421(2):167-72. doi: 10.1016/j.virol.2011.09.024. Epub 2011 Oct 20. Virology. 2011. PMID: 22018782 Free PMC article.
-
Murine gammaherpesvirus 68 contains two functional lytic origins of replication.J Virol. 2007 Jul;81(13):7300-5. doi: 10.1128/JVI.02406-06. Epub 2007 Apr 18. J Virol. 2007. PMID: 17442722 Free PMC article.
-
Comparative Analysis of Gammaherpesvirus Circular RNA Repertoires: Conserved and Unique Viral Circular RNAs.J Virol. 2019 Mar 5;93(6):e01952-18. doi: 10.1128/JVI.01952-18. Print 2019 Mar 15. J Virol. 2019. PMID: 30567979 Free PMC article.
-
ORF30 and ORF34 are essential for expression of late genes in murine gammaherpesvirus 68.J Virol. 2009 Mar;83(5):2265-73. doi: 10.1128/JVI.01785-08. Epub 2008 Dec 17. J Virol. 2009. PMID: 19091863 Free PMC article.
References
-
- Adler, H., M. Messerle, and U. H. Koszinowski. 2003. Cloning of herpesviral genomes as bacterial artificial chromosomes. Rev. Med. Virol. 13:111-121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
