

doi: 10.1128/JVI.78.17.9568-9572.2004.
Model suggesting that replication of influenza virus is regulated by stabilization of replicative intermediates
Affiliations
- PMID: 15308750
- PMCID: PMC506943
- DOI: 10.1128/JVI.78.17.9568-9572.2004
Item in Clipboard
Model suggesting that replication of influenza virus is regulated by stabilization of replicative intermediates
J Virol.
2004 Sep.
Abstract
The RNA-dependent RNA polymerase of influenza A virus is responsible for both transcription and replication of negative-sense viral RNA. It is thought that a "switching" mechanism regulates the transition between these activities. We demonstrate that, in the presence of preexisting viral RNA polymerase and nucleoprotein (NP), influenza A virus synthesizes both mRNA (transcription) and cRNA (replication) early in infection. We suggest that there may be no switch regulating the initiation of RNA synthesis and present a model suggesting that nascent cRNA is degraded by host cell nucleases unless it is stabilized by newly synthesized viral RNA polymerase and NP.
Figures
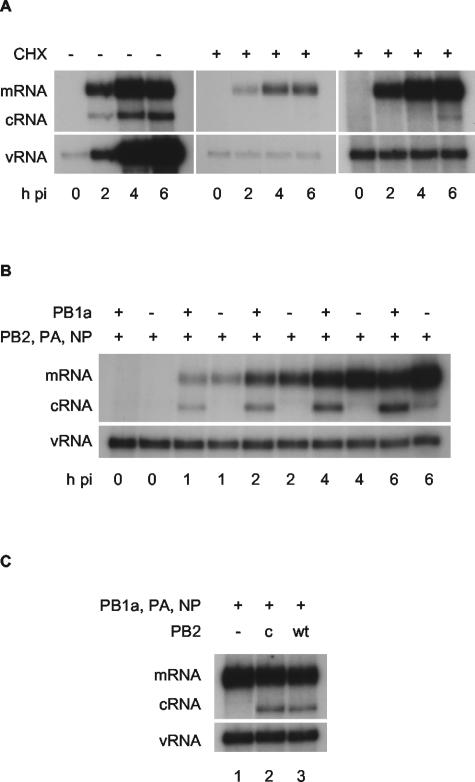
Cycloheximide-mediated inhibition of influenza virus A/WSN/33 cRNA synthesis can be rescued by the expression of PB1, PB2, PA, and NP. Viral RNA species were analyzed by NA gene-specific primer extension assays. mRNA is a broad heterogeneous band 9 to 17 nucleotides longer than cRNA due to its 5′ cap-snatched sequence. (A) Time course of viral infection in the absence (−) or presence (+) of 100 μg of cycloheximide (CHX)/ml. The left-hand and middle panels are equally exposed, with a longer exposure of the middle panel at right showing minor breakthrough of cycloheximide inhibition of cRNA synthesis at 6 h postinfection. (B) Time course of viral infection in the presence of 100 μg of cycloheximide/ml after prior (12 to 14 h) transfection of expression plasmids expressing viral proteins as indicated. (C) cRNA rescue with a cap binding mutant of PB2. 293T cells were transfected with plasmids expressing viral proteins (+) or empty plasmid vector (−), as indicated, 12 to 14 h prior to infection by A/WSN/33 virus in the presence of 100 μg of cycloheximide/ml. RNA was harvested at 2 h postinfection. PB1a, PB1-D445A/D446A; PB2c, PB2-F404A; wt, wild type; h pi, hours postinfection.
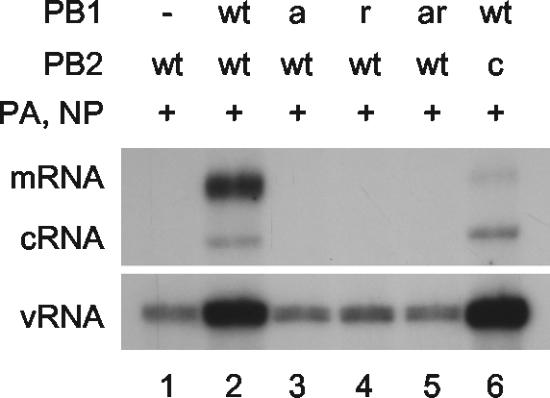
Analysis of the in vivo RNA synthesis activity of polymerase mutants used in the study. Plasmids expressing viral proteins (+) or empty plasmid vector (−), as indicated, were transfected into 293T cells together with a plasmid (pPOLI-NA-RT) (9) directing polymerase I-driven transcription of NA vRNA as a template for RNA synthesis. Total RNA was isolated at 15 h posttransfection and analyzed by NA gene-specific primer extension. vRNA detected in the absence of PB1 (lane 1) represents polymerase I-derived template RNA. wt, wild type; PB1a, PB1-D445A/D446A; PB1r, PB1-Y559A; PB1ar, PB1-D445A/D446A/Y559A; PB2c, PB2-F404A.
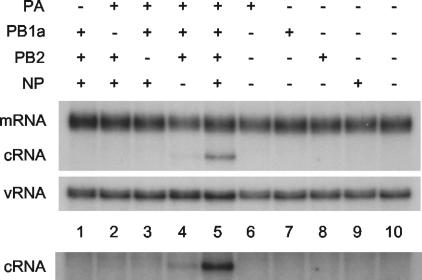
RNA polymerase and NP are essential for rescuing cRNA. 293T cells were transfected with plasmids expressing viral proteins (+) or empty plasmid vector (−), as indicated, 12 to 14 h prior to infection with A/WSN/33 virus in the presence of 100 μg of cycloheximide/ml. RNA was harvested at 2 h postinfection and the viral RNA species were analyzed by NA gene-specific primer extension assays. A longer exposure of the cRNA-specific bands is shown below to emphasise the detection of cRNA in lane 4. PB1a, PB1-D445A/D446A.
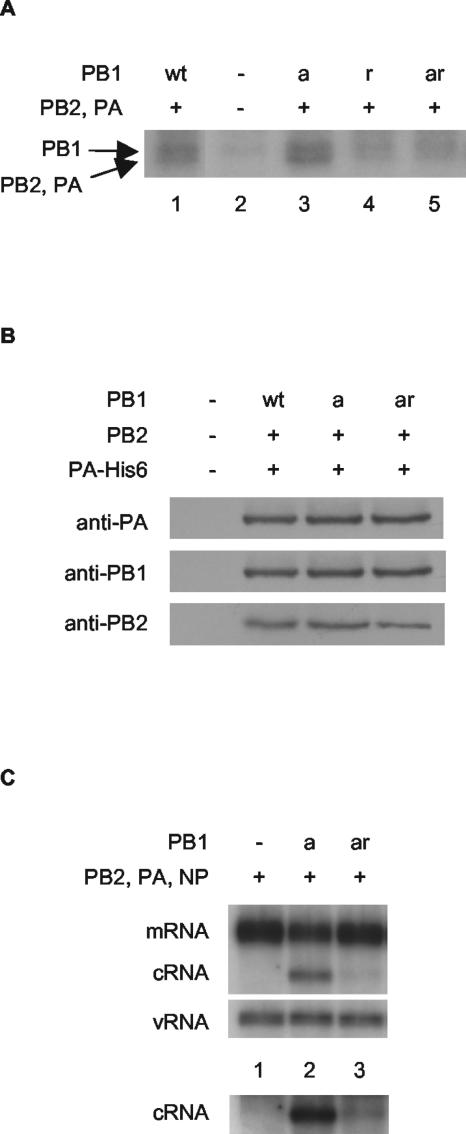
cRNA rescue is reduced by a mutation inhibiting the cRNA promoter-binding activity of PB1. (A) Substitution of tyrosine at residue 559 in PB1 or PB1a with alanine inhibits photochemical cross-linking of the polymerase complex to labeled cRNA. Partially purified His-tagged polymerase from cells transfected with plasmids expressing wild-type or mutant PB1, PB2, and PA-His6 (8), or empty vectors, as indicated, were cross-linked with a 32P-labeled RNA probe corresponding to the 3′-end cRNA in the presence of an excess unlabeled 5′-end cRNA by UV irradiation (8). The products were analyzed by 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and autoradiography. The positions of the PB1 and comigrating PB2 and PA cross-linked bands are shown. (B) The Y559A mutation in PB1 does not inhibit polymerase complex formation. Partially purified His-tagged polymerase from cells transfected with plasmids expressing wild-type or mutant PB1, PB2, and PA-His6 (8), or empty vectors, as indicated, was subjected to Western blotting with antibodies raised against the individual subunits as shown. (C) PB1-Y559A severely restricts cRNA rescue. 293T cells were transfected with plasmids expressing viral proteins (+) or empty plasmid vector (−), as indicated, 12 to 14 h prior to infection by A/WSN/33 virus in the presence of 100 μg of cycloheximide/ml. RNA was harvested at 2 h postinfection, and viral RNA species were analyzed by NA gene-specific primer extension assays. A longer exposure of the cRNA-specific bands is shown below to emphasize the differences in cRNA levels. wt, wild type; PB1a, PB1-D445A/D446A; PB1r, PB1-Y559A; PB1ar, PB1-D445A/D446A/Y559A.
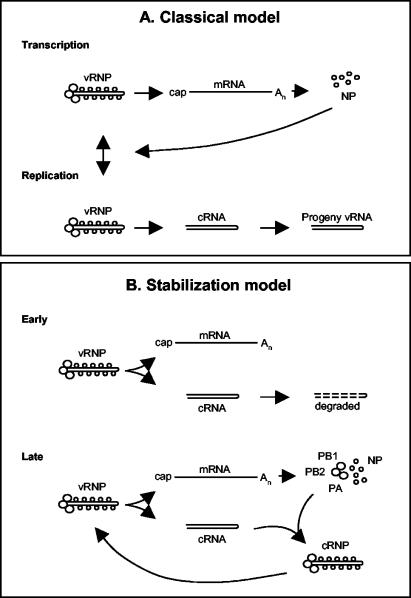
Models for switching between influenza A viral transcription and replication. (A) Classical model. Early in infection, vRNA is transcribed to mRNA (primary transcription). Expression of NP switches primary transcription to replication (synthesis of cRNA and vRNA) and subsequent secondary transcription. (B) Proposed “stabilization” model. Early in infection, mRNA and cRNA are synthesized, but cRNA is degraded. Later in infection, once PB1, PB2, PA, and NP are synthesized, cRNA is stabilized as a cRNP complex that can be replicated to vRNA. See the text for further details.
References
-
- Barr, J. N., S. P. J. Whelan, and G. W. Wertz. 2002. Transcriptional control of the RNA-dependent RNA polymerase of vesicular stomatitis virus. Biochim. Biophys. Acta 1577:337-353. - PubMed
-
- Barrett, T., A. J. Wolstenholme, and B. W. J. Mahy. 1979. Transcription and replication of influenza virus RNA. Virology 98:211-225. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
