

Transcriptome profiling of the response of Arabidopsis suspension culture cells to Suc starvation
- PMID: 15310832
- PMCID: PMC520801
- DOI: 10.1104/pp.104.044362
Transcriptome profiling of the response of Arabidopsis suspension culture cells to Suc starvation
Abstract
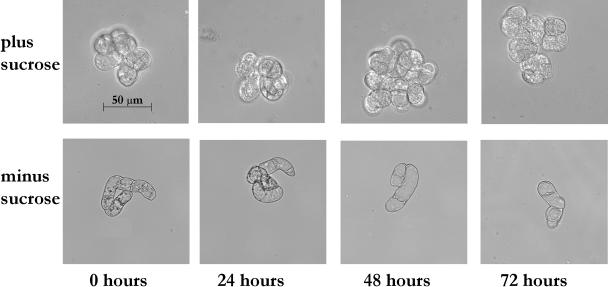
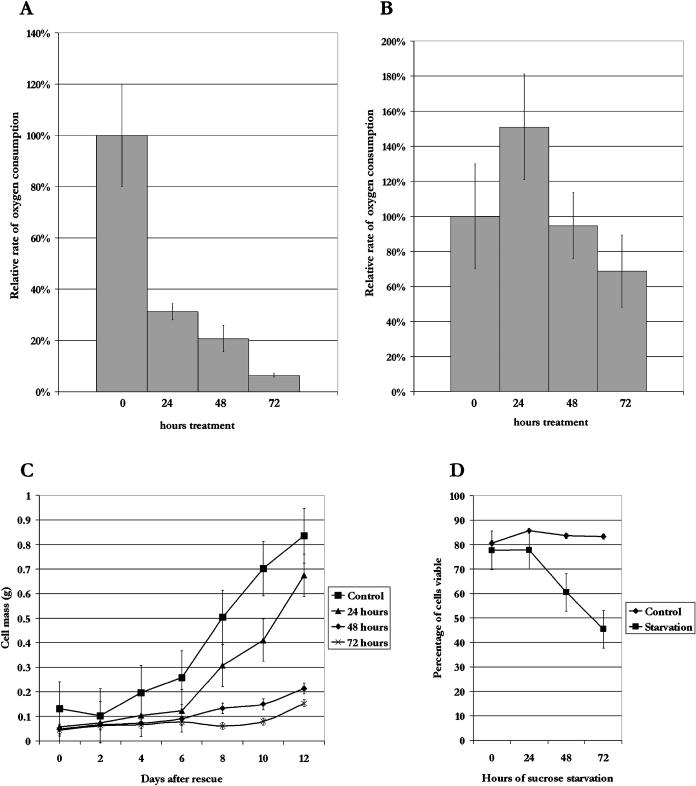
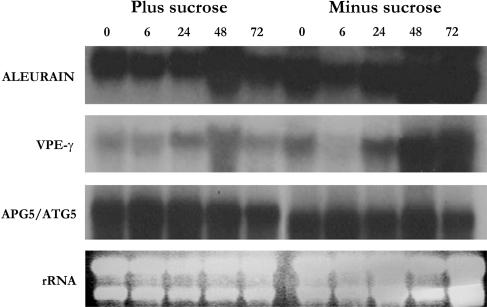
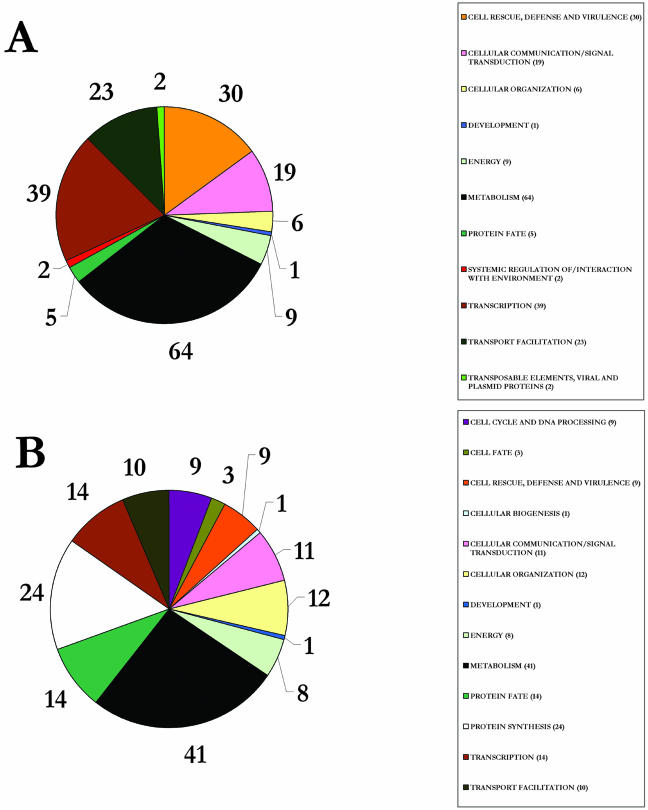
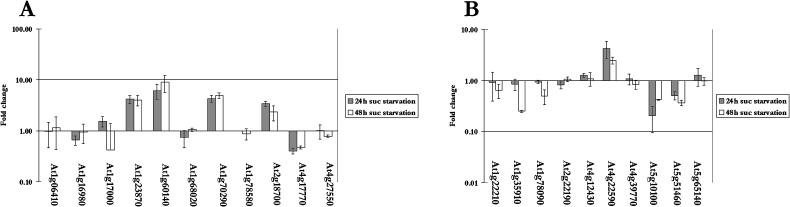
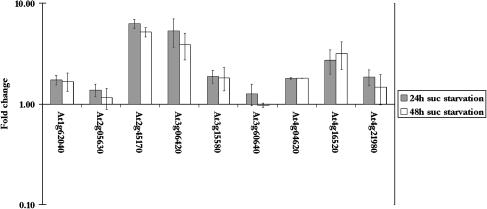
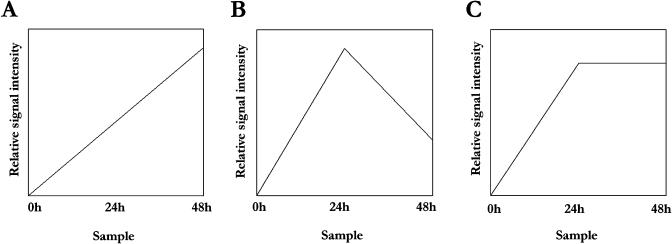
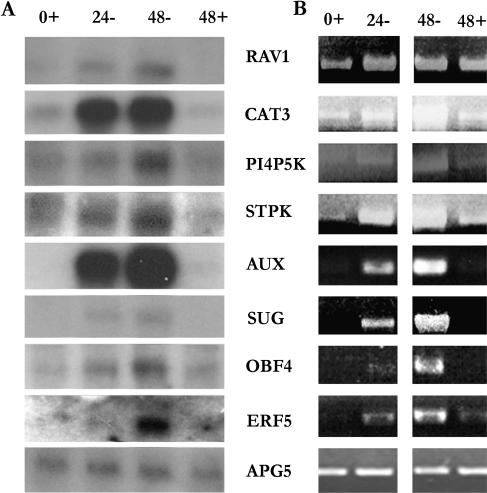
Upon encountering nutrient stress conditions, plant cells undergo extensive metabolic changes and induce nutrient recycling pathways for their continued survival. The role of nutrient mobilization in the response of Arabidopsis suspension cells to Suc starvation was examined. Vacuolar autophagy was induced within 24 h of starvation, with increased expression of vacuolar proteases that are likely to be required for degradation of cytoplasmic components delivered to the vacuole, and thus for nutrient recycling. After 48 h of starvation, culture viability began to decrease, and substantial cell death was evident by 72 h. To provide further insight into the pathways required for survival during Suc deficit, transcriptional profiling during Suc starvation was performed using the ATH1 GeneChip array containing 22,810 probe sets. A significant increase in transcript levels was observed for 343 genes within 48 h of starvation, indicating a response to nutrient stress that utilizes the recycling of cellular components and nutrient scavenging for maintaining cell function, the protection of the cell from death through activation of various defense and stress response pathways, and regulation of these processes by specific protein kinases and transcription factors. These physiological and molecular data support a model in which plant cells initiate a coordinated response of nutrient mobilization at the onset of Suc depletion that is able to maintain cell viability for up to 48 h. After this point, genes potentially involved in cell death increase in expression, whereas those functioning in translation and replication decrease, leading to a decrease in culture viability and activation of cell death programs.
Figures
References
-
- Aubert S, Gout E, Bligny R, Marty-Mazars D, Barrieu F, Alabouvette J, Marty F, Douce R (1996) Ultrastructural and biochemical characterization of autophagy in higher plant cells subjected to carbon deprivation: control by the supply of mitochondria with respiratory substrates. J Cell Biol 133: 1251–1263 - PMC - PubMed
-
- Bassham DC (2002) Golgi-independent trafficking of macromolecules to the plant vacuole. Adv Bot Res 38: 65–92
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
