

Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato
- PMID: 15310835
- PMCID: PMC520773
- DOI: 10.1104/pp.104.048694
Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato
Abstract
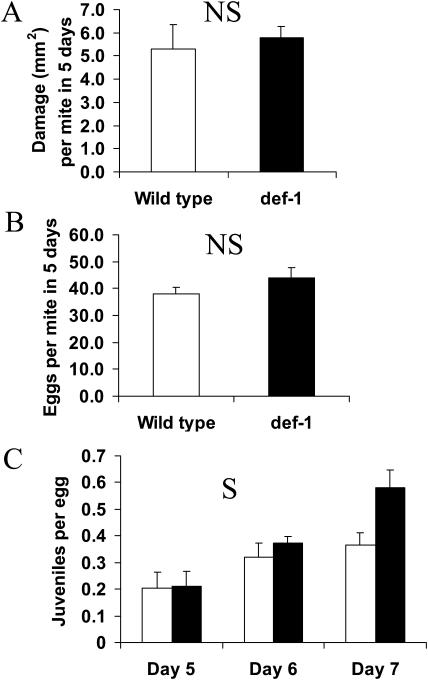
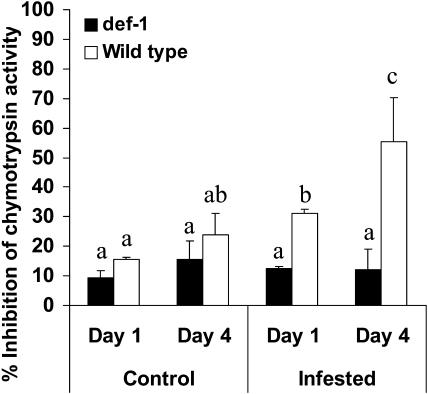
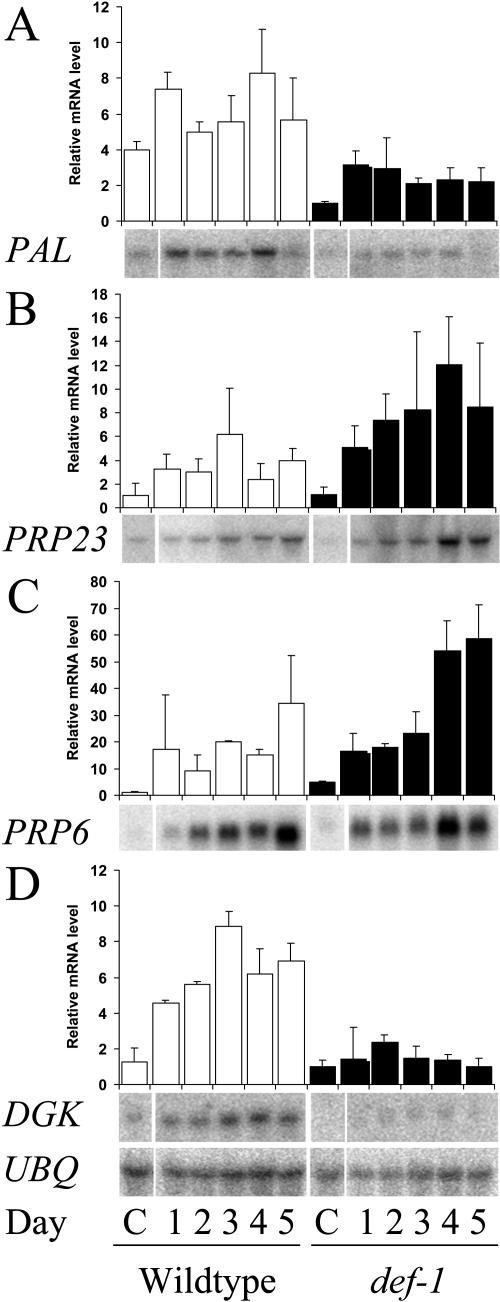
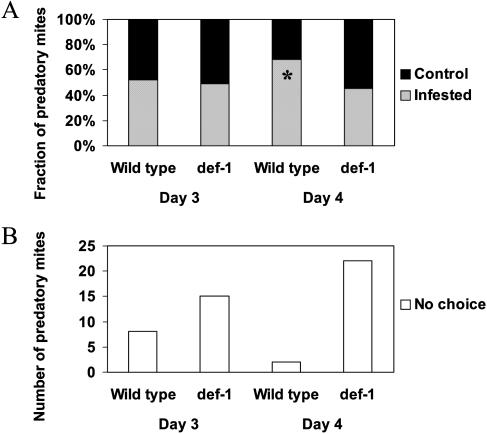
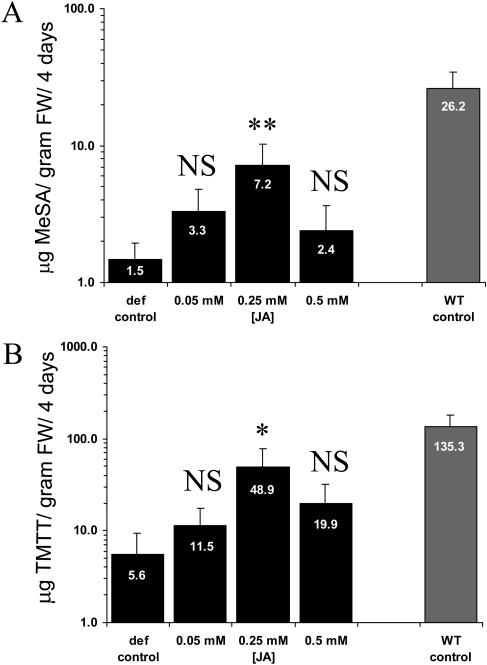
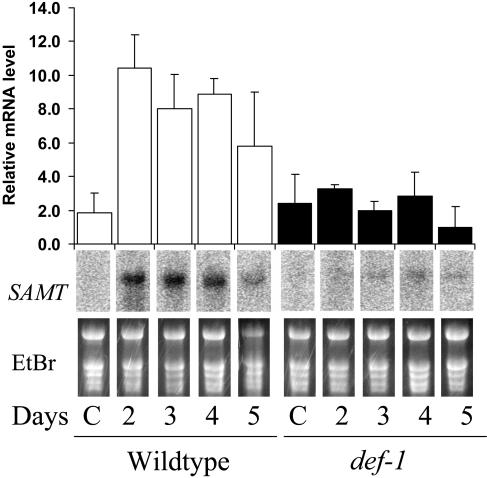
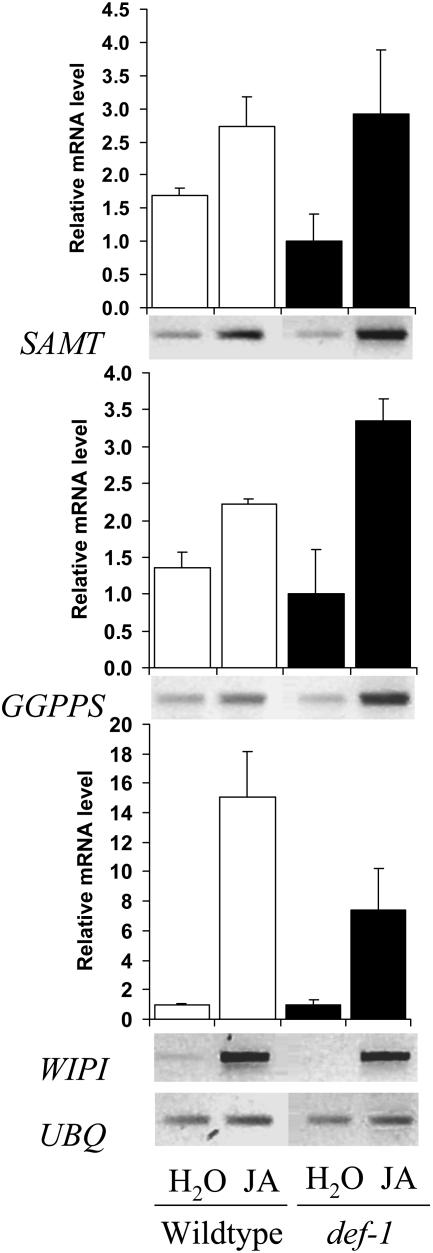
The tomato (Lycopersicon esculentum) mutant def-1, which is deficient in induced jasmonic acid (JA) accumulation upon wounding or herbivory, was used to study the role of JA in the direct and indirect defense responses to phytophagous mites (Tetranychus urticae). In contrast to earlier reports, spider mites laid as many eggs and caused as much damage on def-1 as on wild-type plants, even though def-1 lacked induction of proteinase inhibitor activity. However, the hatching-rate of eggs on def-1 was significantly higher, suggesting that JA-dependent direct defenses enhanced egg mortality or increased the time needed for embryonic development. As to gene expression, def-1 had lower levels of JA-related transcripts but higher levels of salicylic acid (SA) related transcripts after 1 d of spider mite infestation. Furthermore, the indirect defense response was absent in def-1, since the five typical spider mite-induced tomato-volatiles (methyl salicylate [MeSA], 4,8,12-trimethyltrideca-1,3,7,11-tetraene [TMTT], linalool, trans-nerolidol, and trans-beta-ocimene) were not induced and the predatory mite Phytoseiulus persimilis did not discriminate between infested and uninfested def-1 tomatoes as it did with wild-type tomatoes. Similarly, the expression of the MeSA biosynthetic gene salicylic acid methyltransferase (SAMT) was induced by spider mites in wild type but not in def-1. Exogenous application of JA to def-1 induced the accumulation of SAMT and putative geranylgeranyl diphosphate synthase transcripts and restored MeSA- and TMTT-emission upon herbivory. JA is therefore necessary to induce the enzymatic conversion of SA into MeSA. We conclude that JA is essential for establishing the spider mite-induced indirect defense response in tomato.
Figures
References
-
- Arimura G, Ozawa R, Nishioka T, Boland W, Koch T, Kuhnemann F, Takabayashi J (2002) Herbivore-induced volatiles induce emission of ethylene in neighboring lima bean plants. Plant J 29: 87–98 - PubMed
-
- Ashouri A, Overney S, Michaud D, Cloutier C (1998) Fitness and feeding are affected in the two spotted stinkbug, Perillus bioculatus, by the cysteine proteinase inhibitor, oryzacystatin I. Arch Insect Biochem Physiol 38: 74–83
-
- Baldwin IT, Preston CA (1999) The eco-physiological complexity of plant responses to insect herbivores. Planta 208: 137–145
-
- Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Statist Soc B 57: 289–300
-
- Boland W, Gabler A, Gilbert M, Feng Z (1998) Biosynthesis of C11 and C16 homoterpenes in higher plants; stereochemistry of the C-C-bond cleavage reaction. Tetrahedron 54: 14725–14736
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
