

Internal ribosome entry site structural motifs conserved among mammalian fibroblast growth factor 1 alternatively spliced mRNAs
- PMID: 15314170
- PMCID: PMC507008
- DOI: 10.1128/MCB.24.17.7622-7635.2004
Internal ribosome entry site structural motifs conserved among mammalian fibroblast growth factor 1 alternatively spliced mRNAs
Abstract
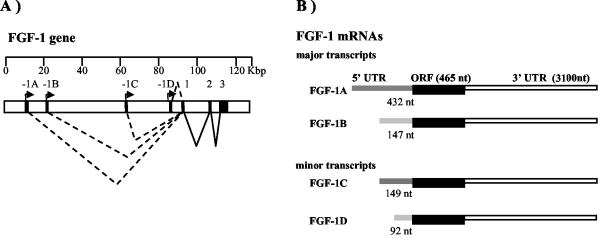
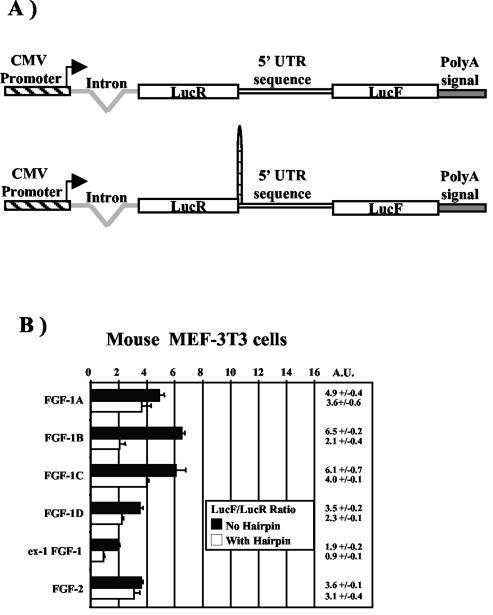
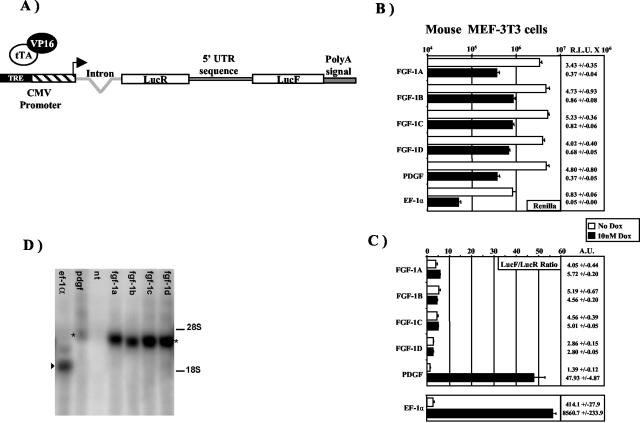
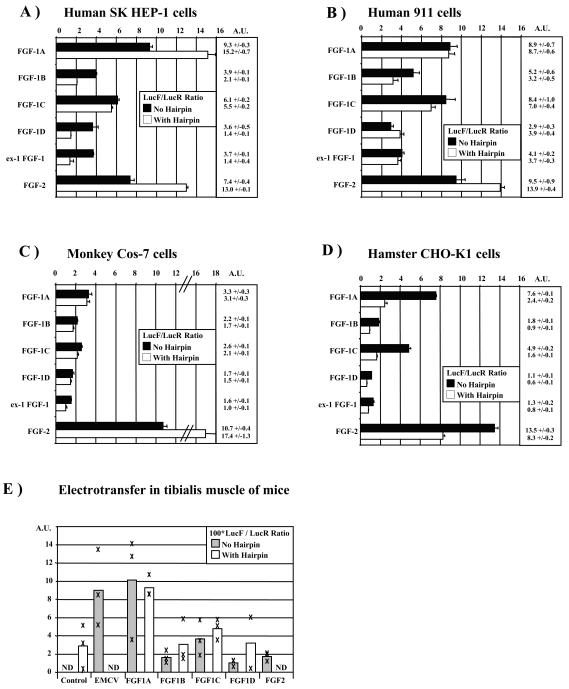
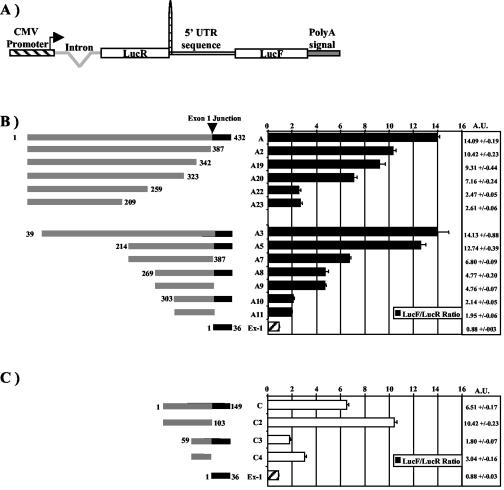
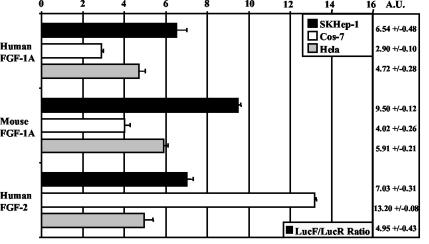
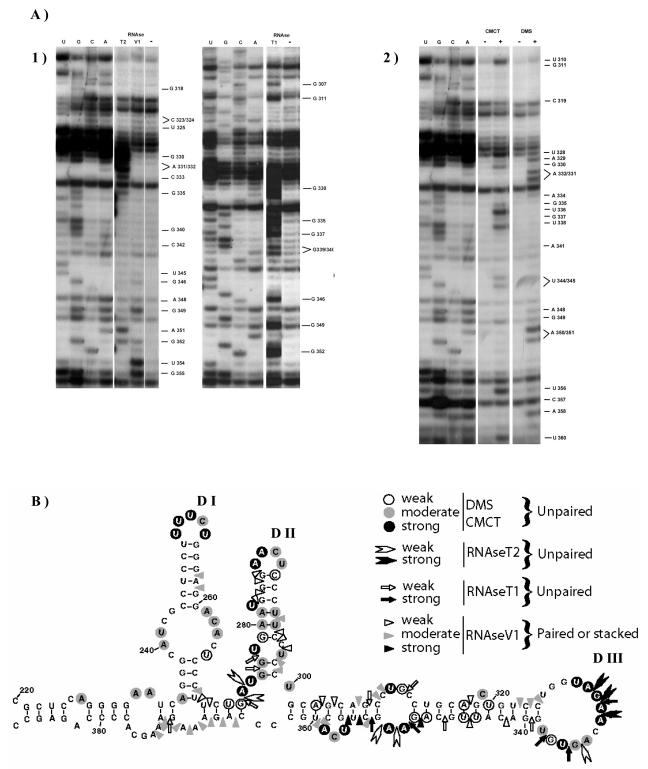
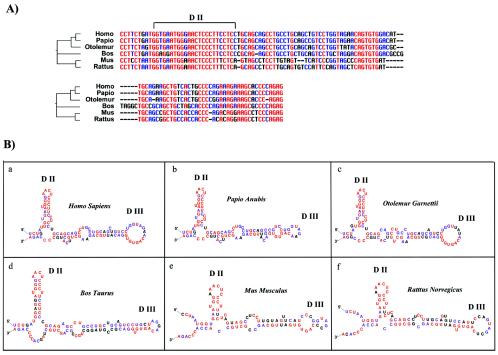
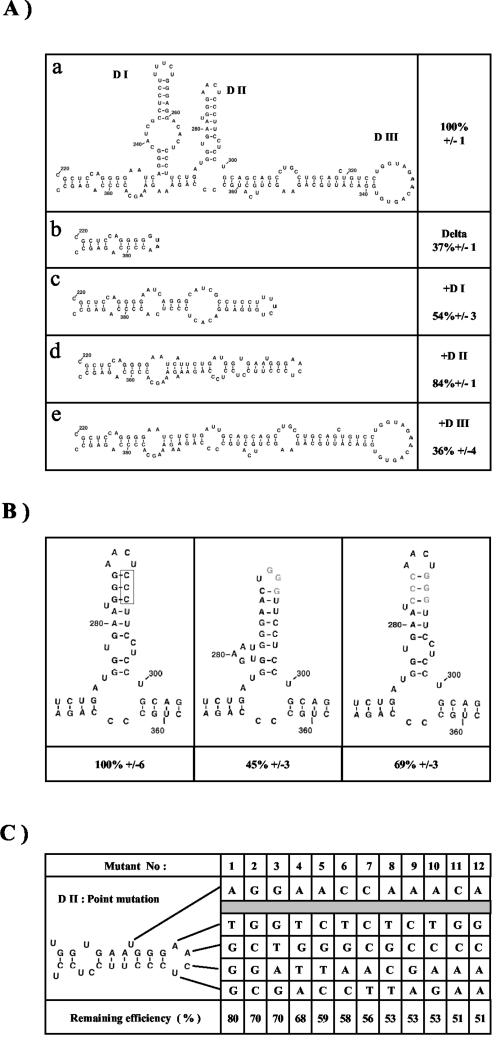
Fibroblast growth factor 1 (FGF-1) is a powerful angiogenic factor whose gene structure contains four promoters, giving rise to a process of alternative splicing resulting in four mRNAs with alternative 5' untranslated regions (5' UTRs). Here we have identified, by using double luciferase bicistronic vectors, the presence of internal ribosome entry sites (IRESs) in the human FGF-1 5' UTRs, particularly in leaders A and C, with distinct activities in mammalian cells. DNA electrotransfer in mouse muscle revealed that the IRES present in the FGF-1 leader A has a high activity in vivo. We have developed a new regulatable TET OFF bicistronic system, which allowed us to rule out the possibility of any cryptic promoter in the FGF-1 leaders. FGF-1 IRESs A and C, which were mapped in fragments of 118 and 103 nucleotides, respectively, are flexible in regard to the position of the initiation codon, making them interesting from a biotechnological point of view. Furthermore, we show that FGF-1 IRESs A of murine and human origins show similar IRES activity profiles. Enzymatic and chemical probing of the FGF-1 IRES A RNA revealed a structural domain conserved among mammals at both the nucleotide sequence and RNA structure levels. The functional role of this structural motif has been demonstrated by point mutagenesis, including compensatory mutations. These data favor an important role of IRESs in the control of FGF-1 expression and provide a new IRES structural motif that could help IRES prediction in 5' UTR databases.
Copyright 2004 American Society for Microbiology
Figures
Similar articles
-
Efficient gene transfer in skeletal muscle with AAV-derived bicistronic vector using the FGF-1 IRES.Gene Ther. 2008 Aug;15(15):1090-8. doi: 10.1038/gt.2008.49. Epub 2008 Mar 27. Gene Ther. 2008. PMID: 18369321
-
Testosterone regulates FGF-2 expression during testis maturation by an IRES-dependent translational mechanism.FASEB J. 2006 Mar;20(3):476-8. doi: 10.1096/fj.04-3314fje. Epub 2006 Jan 19. FASEB J. 2006. PMID: 16423876
-
Long term expression of bicistronic vector driven by the FGF-1 IRES in mouse muscle.BMC Biotechnol. 2007 Oct 28;7:74. doi: 10.1186/1472-6750-7-74. BMC Biotechnol. 2007. PMID: 17963525 Free PMC article.
-
An atypical IRES within the 5' UTR of a dicistrovirus genome.Virus Res. 2009 Feb;139(2):157-65. doi: 10.1016/j.virusres.2008.07.017. Epub 2008 Sep 11. Virus Res. 2009. PMID: 18755228 Review.
-
Ribosomal Chamber Music: Toward an Understanding of IRES Mechanisms.Trends Biochem Sci. 2017 Aug;42(8):655-668. doi: 10.1016/j.tibs.2017.06.002. Epub 2017 Jul 3. Trends Biochem Sci. 2017. PMID: 28684008 Review.
Cited by
-
A search for structurally similar cellular internal ribosome entry sites.Nucleic Acids Res. 2007;35(14):4664-77. doi: 10.1093/nar/gkm483. Epub 2007 Jun 25. Nucleic Acids Res. 2007. PMID: 17591613 Free PMC article.
-
IRES-mediated translation of cofilin regulates axonal growth cone extension and turning.EMBO J. 2018 Mar 1;37(5):e95266. doi: 10.15252/embj.201695266. Epub 2018 Feb 12. EMBO J. 2018. PMID: 29440227 Free PMC article.
-
Dysregulating IRES-dependent translation contributes to overexpression of oncogenic Aurora A Kinase.Mol Cancer Res. 2013 Aug;11(8):887-900. doi: 10.1158/1541-7786.MCR-12-0707. Epub 2013 May 9. Mol Cancer Res. 2013. PMID: 23661421 Free PMC article.
-
Internal ribosome entry site-based vectors for combined gene therapy.World J Exp Med. 2015 Feb 20;5(1):11-20. doi: 10.5493/wjem.v5.i1.11. eCollection 2015 Feb 20. World J Exp Med. 2015. PMID: 25699230 Free PMC article. Review.
-
Cell Fate Control by Translation: mRNA Translation Initiation as a Therapeutic Target for Cancer Development and Stem Cell Fate Control.Biomolecules. 2019 Oct 29;9(11):665. doi: 10.3390/biom9110665. Biomolecules. 2019. PMID: 31671902 Free PMC article. Review.
References
-
- Akiri, G., D. Nahari, Y. Finkelstein, S. Y. Le, O. Elroy-Stein, and B. Z. Levi. 1998. Regulation of vascular endothelial growth factor (VEGF) expression is mediated by internal initiation of translation and alternative initiation of transcription. Oncogene 17:227-236. - PubMed
-
- Bernstein, J., O. Sella, S. Y. Le, and O. Elroy-Stein. 1997. PDGF2/c-sis mRNA leader contains a differentiation-linked internal ribosomal entry site (D-IRES). J. Biol. Chem. 272:9356-9362. - PubMed
-
- Bonnal, S., C. Schaeffer, L. Creancier, S. Clamens, H. Moine, A. C. Prats, and S. Vagner. 2003. A single internal ribosome entry site containing a G quartet RNA structure drives fibroblast growth factor 2 gene expression at four alternative translation initiation codons. J. Biol. Chem. 278:39330-39336. - PMC - PubMed
-
- Brunel, C., and P. Romby. 2000. Probing RNA structure and RNA-ligand complexes with chemical probes. Methods Enzymol. 318:3-21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources