

Cell depletion due to diphtheria toxin fragment A after Cre-mediated recombination
- PMID: 15314171
- PMCID: PMC506983
- DOI: 10.1128/MCB.24.17.7636-7642.2004
Cell depletion due to diphtheria toxin fragment A after Cre-mediated recombination
Abstract
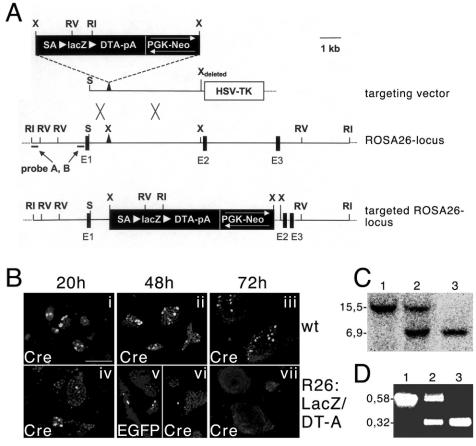
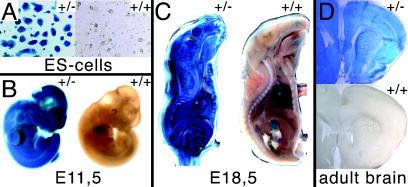
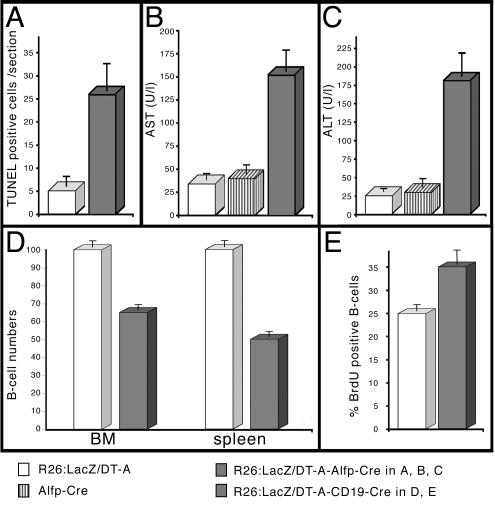
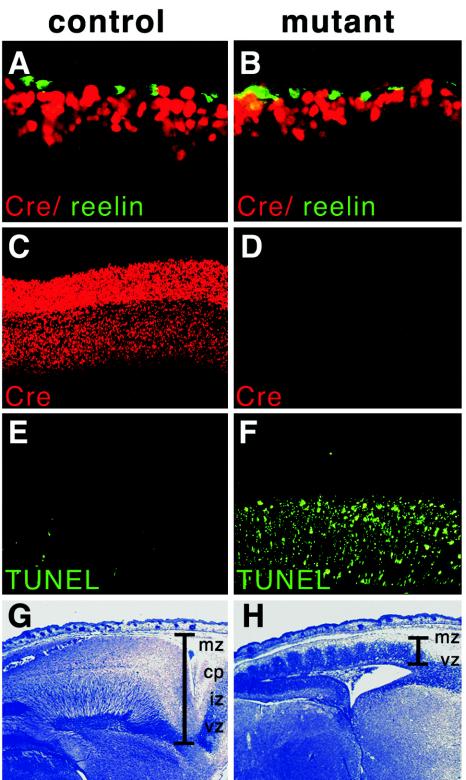
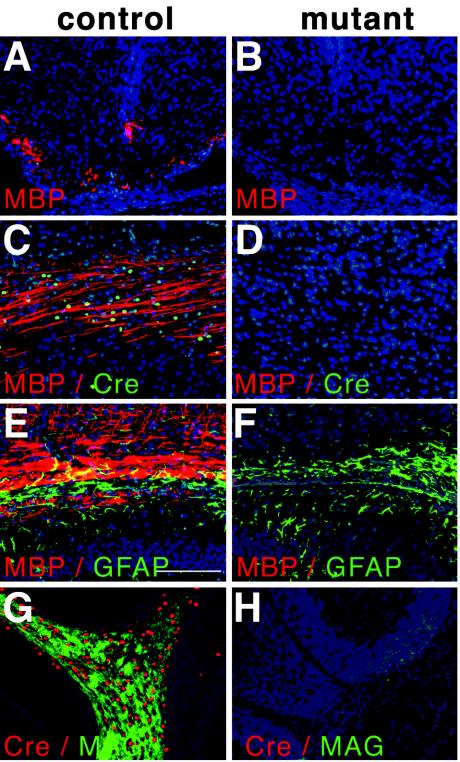
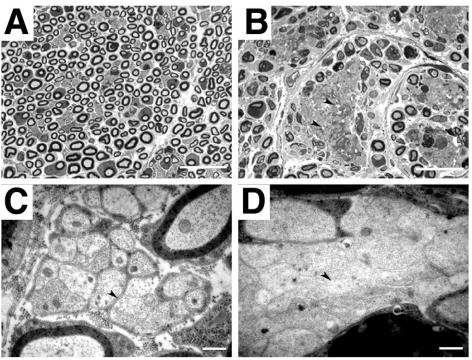
Abnormal cell loss is the common cause of a large number of developmental and degenerative diseases. To model such diseases in transgenic animals, we have developed a line of mice that allows the efficient depletion of virtually any cell type in vivo following somatic Cre-mediated gene recombination. By introducing the diphtheria toxin fragment A (DT-A) gene as a conditional expression construct (floxed lacZ-DT-A) into the ubiquitously expressed ROSA26 locus, we produced a line of mice that would permit cell-specific activation of the toxin gene. Following Cre-mediated recombination under the control of cell-type-specific promoters, lacZ gene expression was efficiently replaced by de novo transcription of the Cre-recombined DT-A gene. We provide proof of this principle, initially for cells of the central nervous system (pyramidal neurons and oligodendrocytes), the immune system (B cells), and liver tissue (hepatocytes), that the conditional expression of DT-A is functional in vivo, resulting in the generation of novel degenerative disease models.
Copyright 2004 American Society for Microbiology
Figures
References
-
- Awatramani, R., P. Soriano, J. J. Mai, and S. Dymecki. 2001. An Flp indicator mouse expressing alkaline phosphatase from the ROSA26 locus. Nat. Genet. 29:257-259. - PubMed
-
- Bartholoma, A., and K. A. Nave. 1994. NEX-1: a novel brain-specific helix-loop-helix protein with autoregulation and sustained expression in mature cortical neurons. Mech. Dev. 48:217-228. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases