

Jun blockade of erythropoiesis: role for repression of GATA-1 by HERP2
- PMID: 15314183
- PMCID: PMC506977
- DOI: 10.1128/MCB.24.17.7779-7794.2004
Jun blockade of erythropoiesis: role for repression of GATA-1 by HERP2
Erratum in
-
Erratum for Elagib et al., Jun Blockade of Erythropoiesis: Role for Repression of GATA-1 by HERP2.Mol Cell Biol. 2015 Jun;35(12):2215. doi: 10.1128/MCB.00374-15. Mol Cell Biol. 2015. PMID: 25987232 Free PMC article. No abstract available.
Abstract
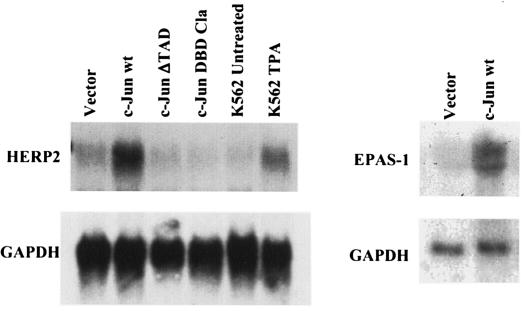
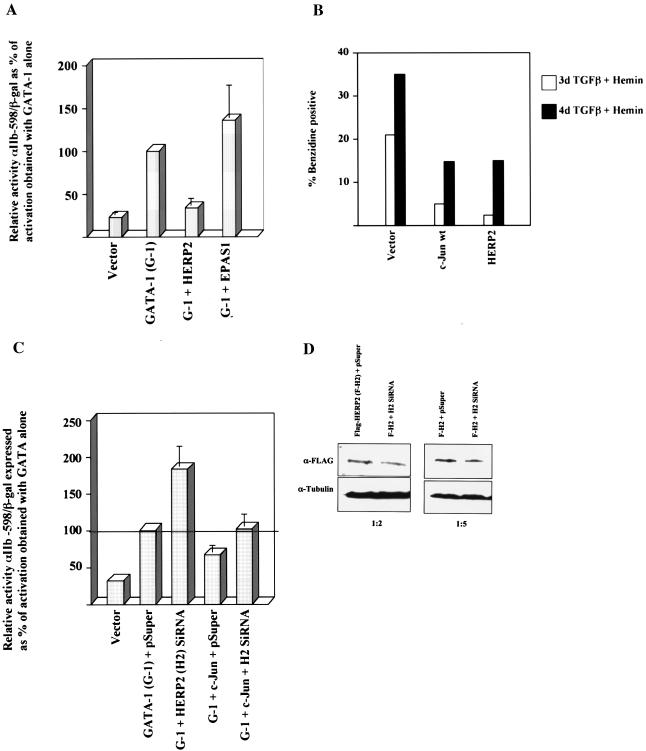
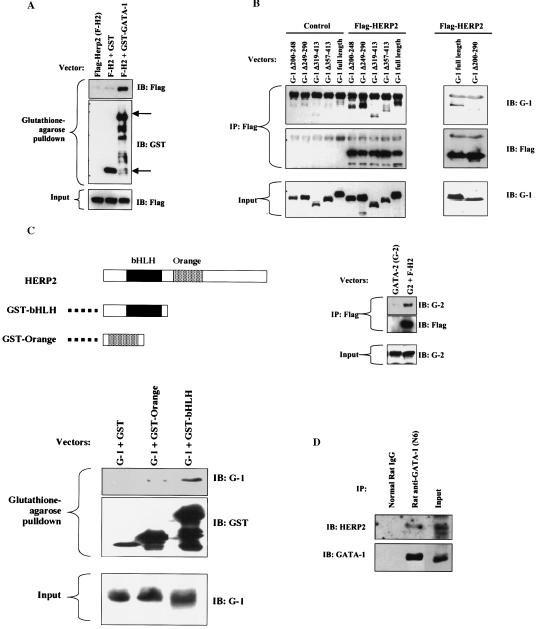
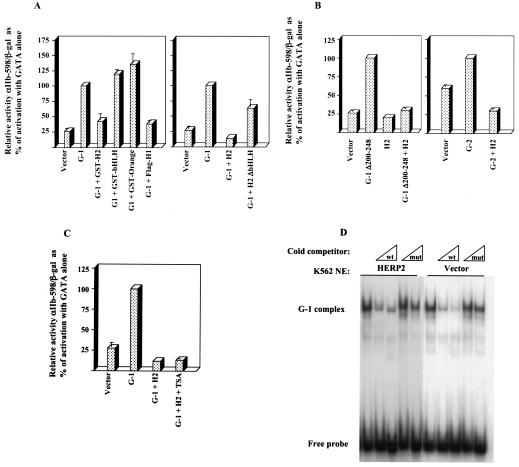
Although Jun upregulation and activation have been established as critical to oncogenesis, the relevant downstream pathways remain incompletely characterized. In this study, we found that c-Jun blocks erythroid differentiation in primary human hematopoietic progenitors and, correspondingly, that Jun factors block transcriptional activation by GATA-1, the central regulator of erythroid differentiation. Mutagenesis of c-Jun suggested that its repression of GATA-1 occurs through a transcriptional mechanism involving activation of downstream genes. We identified the hairy-enhancer-of-split-related factor HERP2 as a novel gene upregulated by c-Jun. HERP2 showed physical interaction with GATA-1 and repressed GATA-1 transcriptional activation. Furthermore, transduction of HERP2 into primary human hematopoietic progenitors inhibited erythroid differentiation. These results thus define a novel regulatory pathway linking the transcription factors c-Jun, HERP2, and GATA-1. Furthermore, these results establish a connection between the Notch signaling pathway, of which the HERP factors are a critical component, and the GATA family, which participates in programming of cellular differentiation.
Copyright 2004 American Society for Microbiology
Figures
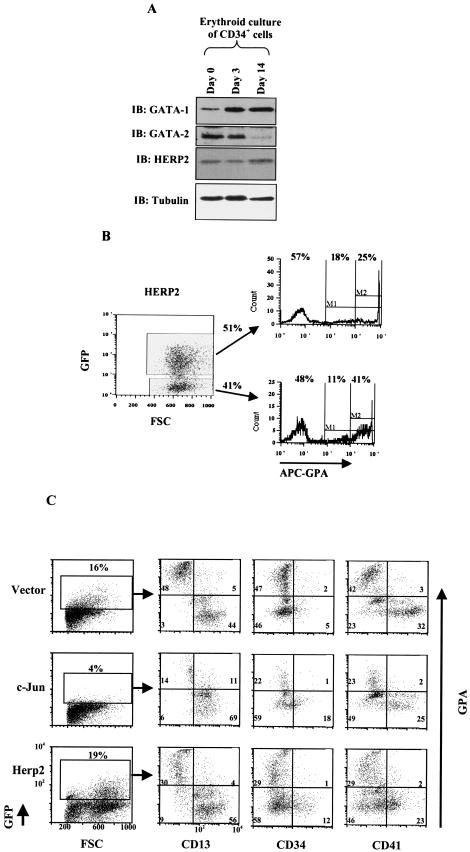
Similar articles
-
Control of erythroid cell production via caspase-mediated cleavage of transcription factor SCL/Tal-1.Cell Death Differ. 2003 Aug;10(8):905-13. doi: 10.1038/sj.cdd.4401255. Cell Death Differ. 2003. PMID: 12867998
-
The regulatory roles of microRNA-146b-5p and its target platelet-derived growth factor receptor α (PDGFRA) in erythropoiesis and megakaryocytopoiesis.J Biol Chem. 2014 Aug 15;289(33):22600-22613. doi: 10.1074/jbc.M114.547380. Epub 2014 Jun 30. J Biol Chem. 2014. PMID: 24982425 Free PMC article.
-
GATA-1 utilizes Ikaros and polycomb repressive complex 2 to suppress Hes1 and to promote erythropoiesis.Mol Cell Biol. 2012 Sep;32(18):3624-38. doi: 10.1128/MCB.00163-12. Epub 2012 Jul 9. Mol Cell Biol. 2012. PMID: 22778136 Free PMC article.
-
Regulation of the transcription factor GATA-1 at the gene and protein level.Cell Mol Life Sci. 2001 Dec;58(14):2008-17. doi: 10.1007/PL00000833. Cell Mol Life Sci. 2001. PMID: 11814053 Free PMC article. Review.
-
Autophagy as a regulatory component of erythropoiesis.Int J Mol Sci. 2015 Feb 13;16(2):4083-94. doi: 10.3390/ijms16024083. Int J Mol Sci. 2015. PMID: 25689426 Free PMC article. Review.
Cited by
-
Notch Stimulates Both Self-Renewal and Lineage Plasticity in a Subset of Murine CD9High Committed Megakaryocytic Progenitors.PLoS One. 2016 Apr 18;11(4):e0153860. doi: 10.1371/journal.pone.0153860. eCollection 2016. PLoS One. 2016. PMID: 27089435 Free PMC article.
-
The leukemia associated nuclear corepressor ETO homologue genes MTG16 and MTGR1 are regulated differently in hematopoietic cells.BMC Mol Biol. 2012 Mar 23;13:11. doi: 10.1186/1471-2199-13-11. BMC Mol Biol. 2012. PMID: 22443175 Free PMC article.
-
Dissection of the transformation of primary human hematopoietic cells by the oncogene NUP98-HOXA9.PLoS One. 2009 Aug 21;4(8):e6719. doi: 10.1371/journal.pone.0006719. PLoS One. 2009. PMID: 19696924 Free PMC article.
-
Iron control of erythroid development by a novel aconitase-associated regulatory pathway.Blood. 2010 Jul 8;116(1):97-108. doi: 10.1182/blood-2009-10-251496. Epub 2010 Apr 20. Blood. 2010. PMID: 20407036 Free PMC article.
-
Neonatal expression of RNA-binding protein IGF2BP3 regulates the human fetal-adult megakaryocyte transition.J Clin Invest. 2017 Jun 1;127(6):2365-2377. doi: 10.1172/JCI88936. Epub 2017 May 8. J Clin Invest. 2017. PMID: 28481226 Free PMC article.
References
-
- Alitalo, R., J. Partanen, L. Pertovaara, E. Holtta, L. Sistonen, L. Andersson, and K. Alitalo. 1990. Increased erythroid potentiating activity/tissue inhibitor of metalloproteinases and jun/fos transcription factor complex characterize tumor promoter-induced megakaryoblastic differentiation of K562 leukemia cells. Blood 75:1974-1982. - PubMed
-
- Bamberger, A. M., K. Milde-Langosch, E. Rossing, C. Goemann, and T. Loning. 2001. Expression pattern of the AP-1 family in endometrial cancer: correlations with cell cycle regulators. J. Cancer Res. Clin. Oncol. 127:545-550. - PubMed
-
- Bauknecht, T., P. Angel, M. Kohler, F. Kommoss, G. Birmelin, A. Pfleiderer, and E. Wagner. 1993. Gene structure and expression analysis of the epidermal growth factor receptor, transforming growth factor-alpha, myc, jun, and metallothionein in human ovarian carcinomas. Classification of malignant phenotypes. Cancer 71:419-429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous