

Spliced-leader RNA trans splicing in a chordate, Oikopleura dioica, with a compact genome
- PMID: 15314184
- PMCID: PMC507004
- DOI: 10.1128/MCB.24.17.7795-7805.2004
Spliced-leader RNA trans splicing in a chordate, Oikopleura dioica, with a compact genome
Abstract
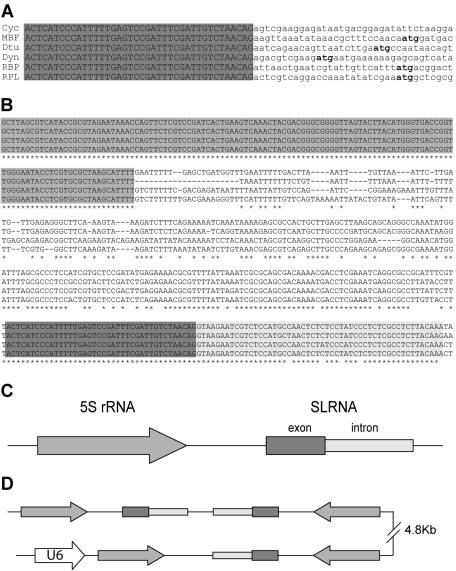
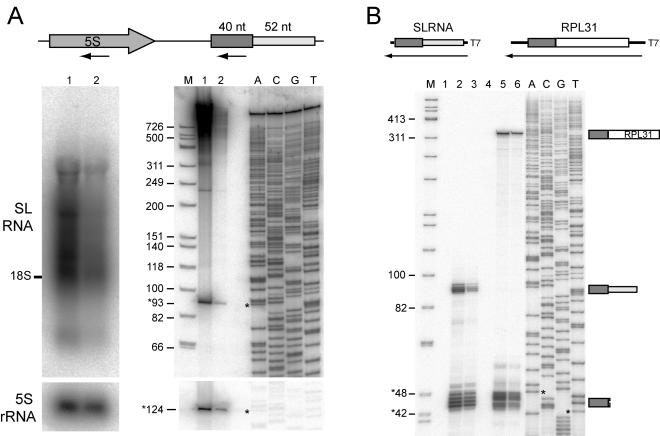
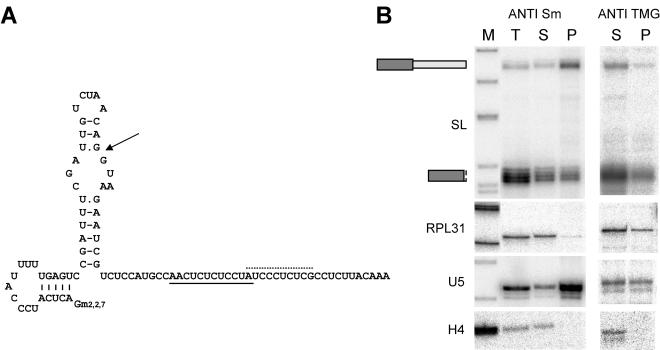
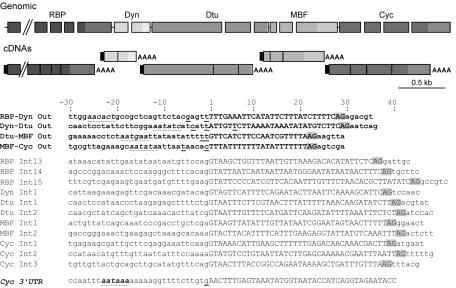
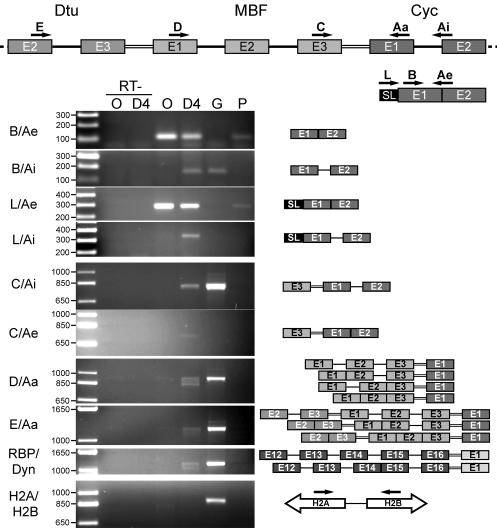
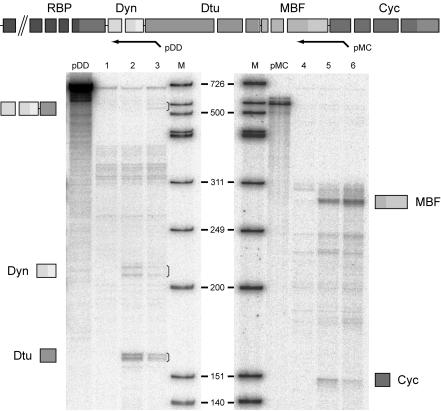
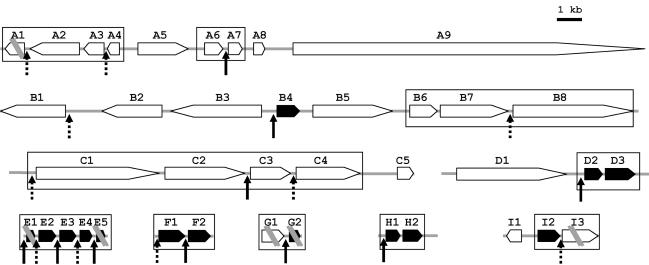
trans splicing of a spliced-leader RNA (SL RNA) to the 5' ends of mRNAs has been shown to have a limited and sporadic distribution among eukaryotes. Within metazoans, only nematodes are known to process polycistronic pre-mRNAs, produced from operon units of transcription, into mature monocistronic mRNAs via an SL RNA trans-splicing mechanism. Here we demonstrate that a chordate with a highly compact genome, Oikopleura dioica, now joins Caenorhabditis elegans in coupling trans splicing with processing of polycistronic transcipts. We identified a single SL RNA which associates with Sm proteins and has a trimethyl guanosine cap structure reminiscent of spliceosomal snRNPs. The same SL RNA, estimated to be trans-spliced to at least 25% of O. dioica mRNAs, is used for the processing of both isolated or first cistrons and downstream cistrons in a polycistronic precursor. Remarkably, intercistronic regions in O. dioica are far more reduced than those in either nematodes or kinetoplastids, implying minimal cis-regulatory elements for coupling of 3'-end formation and trans splicing.
Copyright 2004 American Society for Microbiology
Figures
References
-
- Blumenthal, T. 1995. trans-splicing and polycistronic transcription in Caenorhabditis elegans. Trends Genet. 11:132-136. - PubMed
-
- Campbell, D. A., N. R. Sturm, and M. C. Yu. 2000. Transcription of the kinetoplastid spliced leader RNA gene. Parasitol. Today 16:78-82. - PubMed
-
- Chioda, M., R. Eskeland, and E. M. Thompson. 2002. Histone gene complement, variant expression, and mRNA processing in a urochordate Oikopleura dioica that undergoes extensive polyploidization. Mol. Biol. Evol. 19:2247-2260. - PubMed
-
- Colgan, D. F., and J. L. Manley. 1997. Mechanism and regulation of mRNA polyadenylation. Genes Dev. 11:2755-2766. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous