

The Bvg virulence control system regulates biofilm formation in Bordetella bronchiseptica
- PMID: 15317773
- PMCID: PMC516841
- DOI: 10.1128/JB.186.17.5692-5698.2004
The Bvg virulence control system regulates biofilm formation in Bordetella bronchiseptica
Abstract
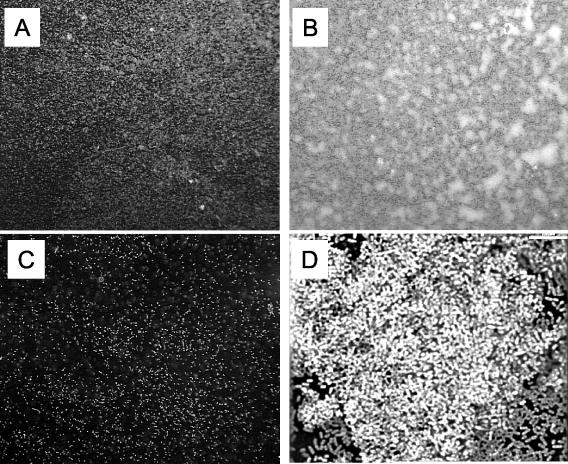
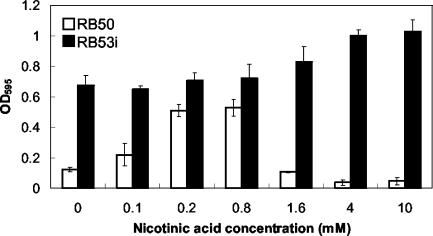
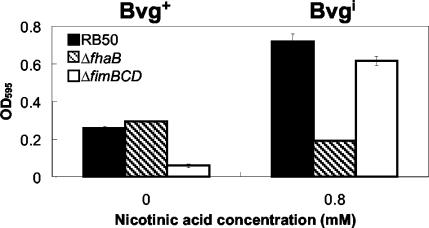
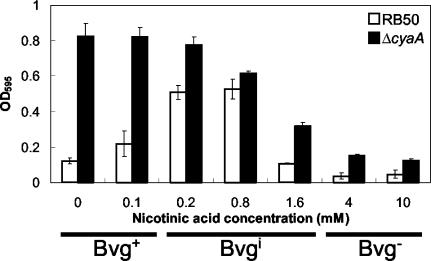
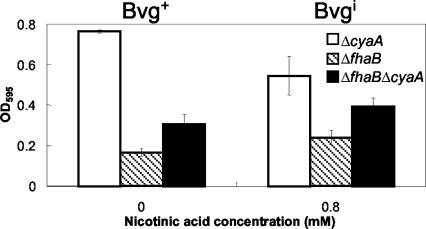
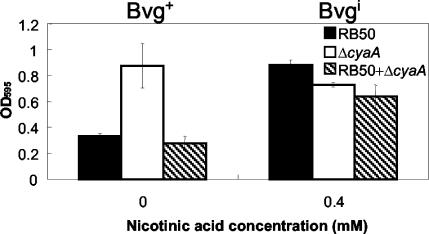
Bordetella species utilize the BvgAS (Bordetella virulence gene) two-component signal transduction system to sense the environment and regulate gene expression among at least three phases: a virulent Bvg+ phase, a nonvirulent Bvg- phase, and an intermediate Bvgi phase. Genes expressed in the Bvg+ phase encode known virulence factors, including adhesins such as filamentous hemagglutinin (FHA) and fimbriae, as well as toxins such as the bifunctional adenylate cyclase/hemolysin (ACY). Previous studies showed that in the Bvgi phase, FHA and fimbriae continue to be expressed, but ACY expression is significantly downregulated. In this report, we determine that Bordetella bronchiseptica can form biofilms in vitro and that the generation of biofilm is maximal in the Bvgi phase. We show that FHA is required for maximal biofilm formation and that fimbriae may also contribute to this phenotype. However, expression of ACY inhibits biofilm formation, most likely via interactions with FHA. Therefore, the coordinated regulation of adhesins and ACY expression leads to maximal biofilm formation in the Bvgi phase in B. bronchiseptica.
Figures

References
-
- Akerley, B. J., P. A. Cotter, and J. F. Miller. 1995. Ectopic expression of the flagellar regulon alters development of the Bordetella-host interaction. Cell 80:611-620. - PubMed
-
- Allison, D. G., B. Ruiz, C. SanJose, A. Jaspe, and P. Gilbert. 1998. Extracellular products as mediators of the formation and detachment of Pseudomonas fluorescens biofilms. FEMS Microbiol. Lett. 167:179-184. - PubMed
-
- Austin, J. W., G. Sanders, W. W. Kay, and S. K. Collinson. 1998. Thin aggregative fimbriae enhance Salmonella enteritidis biofilm formation. FEMS Microbiol. Lett. 162:295-301. - PubMed
-
- Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318-1322. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
