

The N-terminal half-domain of the long form of tRNase Z is required for the RNase 65 activity
- PMID: 15317868
- PMCID: PMC516050
- DOI: 10.1093/nar/gkh774
The N-terminal half-domain of the long form of tRNase Z is required for the RNase 65 activity
Abstract
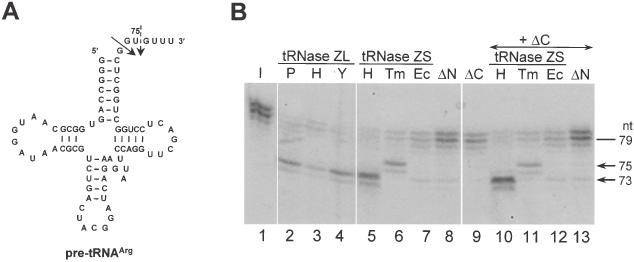
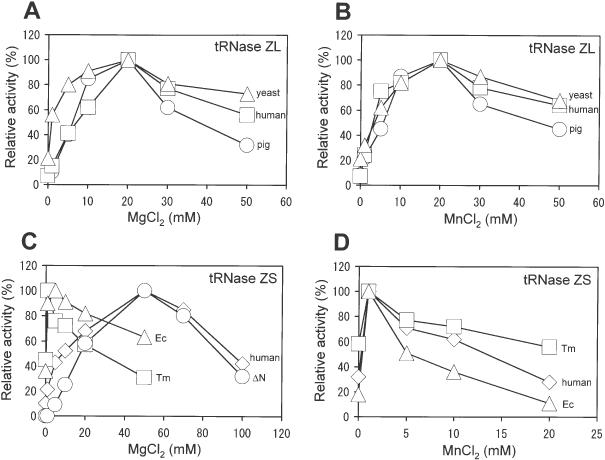
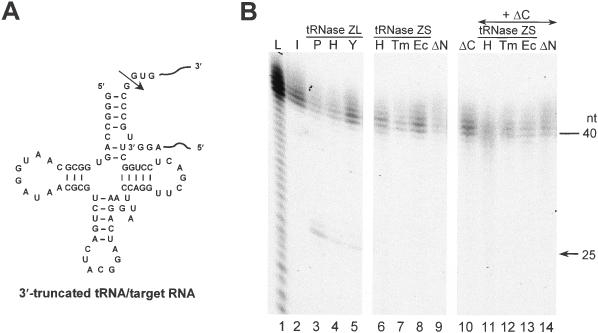
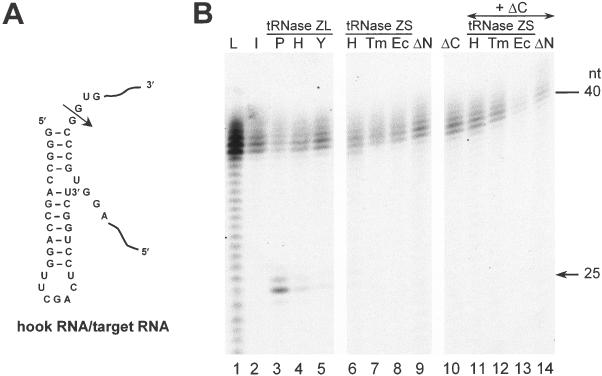
Transfer RNA (tRNA) 3' processing endoribonuclease (tRNase Z) is an enzyme responsible for the removal of a 3' trailer from pre-tRNA. There exists two types of tRNase Z: one is a short form (tRNase ZS) that consists of 300-400 amino acids, and the other is a long form (tRNase ZL) that contains 800-900 amino acids. Here we investigated whether the short and long forms have different preferences for various RNA substrates. We examined three recombinant tRNase ZSs from human, Escherichia coli and Thermotoga maritima, two recombinant tRNase ZLs from human and Saccharomyces cerevisiae, one tRNase ZL from pig liver, and the N- and C-terminal half regions of human tRNase ZL for cleavage of human micro-pre-tRNA(Arg) and the RNase 65 activity. All tRNase ZLs cleaved the micro-pre-tRNA and showed the RNase 65 activity, while all tRNase ZSs and both half regions of human tRNase ZL failed to do so with the exception of the C-terminal half, which barely cleaved the micro-pre-tRNA. We also show that only the long forms of tRNase Z can specifically cleave a target RNA under the direction of a new type of small guide RNA, hook RNA. These results indicate that indeed tRNase ZL and tRNase ZS have different substrate specificities and that the differences are attributed to the N-terminal half-domain of tRNase ZL. Furthermore, the optimal concentrations of NaCl, MgCl2 and MnCl2 differed between tRNase ZSs and tRNase ZLs, and the K(m) values implied that tRNase ZLs interact with pre-tRNA substrates more strongly than tRNase ZSs.
Figures



Similar articles
-
The flexible arm of tRNase Z is not essential for pre-tRNA binding but affects cleavage site selection.J Mol Biol. 2008 Aug 29;381(2):289-99. doi: 10.1016/j.jmb.2008.05.016. Epub 2008 May 15. J Mol Biol. 2008. PMID: 18602113
-
Specific cleavage of target RNAs from HIV-1 with 5' half tRNA by mammalian tRNA 3' processing endoribonuclease.RNA. 1996 Jun;2(6):523-4. RNA. 1996. PMID: 8718682 Free PMC article.
-
Unstructured RNA is a substrate for tRNase Z.Biochemistry. 2006 May 2;45(17):5486-92. doi: 10.1021/bi051972s. Biochemistry. 2006. PMID: 16634630
-
The tRNase Z family of proteins: physiological functions, substrate specificity and structural properties.Biol Chem. 2005 Dec;386(12):1253-64. doi: 10.1515/BC.2005.142. Biol Chem. 2005. PMID: 16336119 Review.
-
The making of tRNAs and more - RNase P and tRNase Z.Prog Mol Biol Transl Sci. 2009;85:319-68. doi: 10.1016/S0079-6603(08)00808-8. Prog Mol Biol Transl Sci. 2009. PMID: 19215776 Review.
Cited by
-
The structure of the flexible arm of Thermotoga maritima tRNase Z differs from those of homologous enzymes.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2007 Aug 1;63(Pt 8):637-41. doi: 10.1107/S1744309107033623. Epub 2007 Jul 21. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2007. PMID: 17671357 Free PMC article.
-
Characterization of TRZ1, a yeast homolog of the human candidate prostate cancer susceptibility gene ELAC2 encoding tRNase Z.BMC Mol Biol. 2005 May 13;6:12. doi: 10.1186/1471-2199-6-12. BMC Mol Biol. 2005. PMID: 15892892 Free PMC article.
-
Elimination of specific miRNAs by naked 14-nt sgRNAs.PLoS One. 2012;7(6):e38496. doi: 10.1371/journal.pone.0038496. Epub 2012 Jun 4. PLoS One. 2012. PMID: 22675569 Free PMC article.
-
Inhibition of HIV-1 gene expression by retroviral vector-mediated small-guide RNAs that direct specific RNA cleavage by tRNase ZL.Nucleic Acids Res. 2005 Jan 12;33(1):235-43. doi: 10.1093/nar/gki164. Print 2005. Nucleic Acids Res. 2005. PMID: 15647506 Free PMC article.
-
Partitioning of the nuclear and mitochondrial tRNA 3'-end processing activities between two different proteins in Schizosaccharomyces pombe.J Biol Chem. 2013 Sep 20;288(38):27415-27422. doi: 10.1074/jbc.M113.501569. Epub 2013 Aug 8. J Biol Chem. 2013. PMID: 23928301 Free PMC article.
References
-
- Tavtigian S.V., Simard,J., Teng,D.H., Abtin,V., Baumgard,M., Beck,A., Camp,N.J., Carillo,A.R., Chen,Y., Dayananth,P. et al. (2001) A candidate prostate cancer susceptibility gene at chromosome 17p. Nature Genet., 27, 172–180. - PubMed
-
- Minagawa A., Takaku,H., Takagi,M. and Nashimoto,M. (2004) A novel endonucleolytic mechanism to generate the CCA 3′ termini of tRNA molecules in Thermotoga maritima. J. Biol. Chem., 279, 15688–15697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
