

A cryptic matrix targeting signal of the yeast ADP/ATP carrier normally inserted by the TIM22 complex is recognized by the TIM23 machinery
- PMID: 15320873
- PMCID: PMC1134685
- DOI: 10.1042/BJ20040650
A cryptic matrix targeting signal of the yeast ADP/ATP carrier normally inserted by the TIM22 complex is recognized by the TIM23 machinery
Abstract
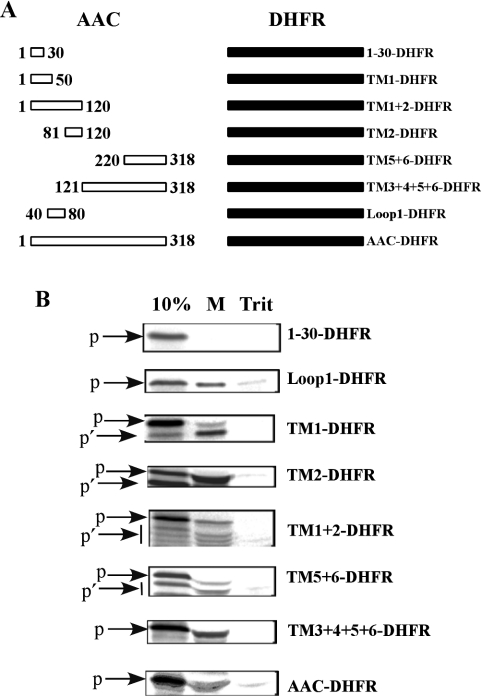
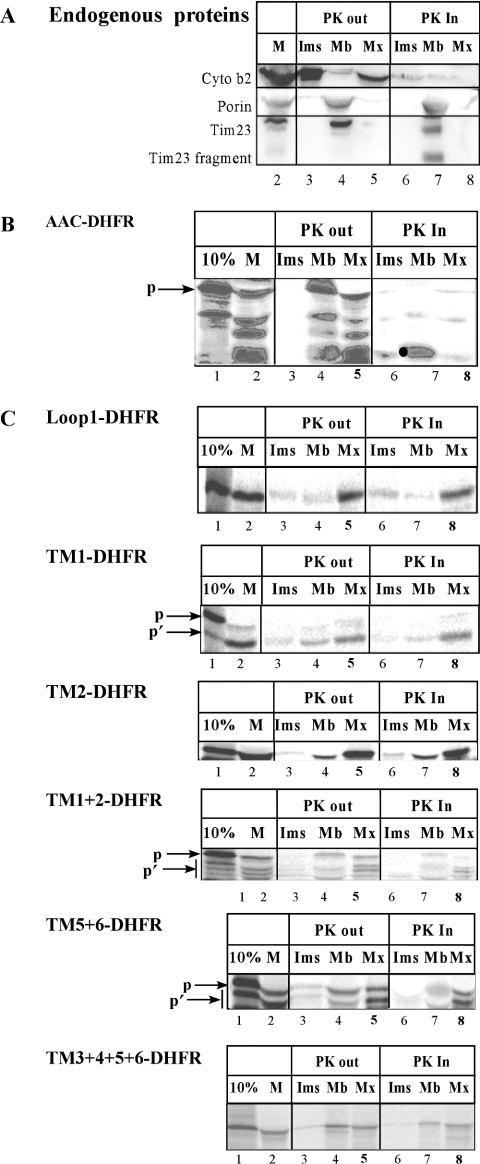
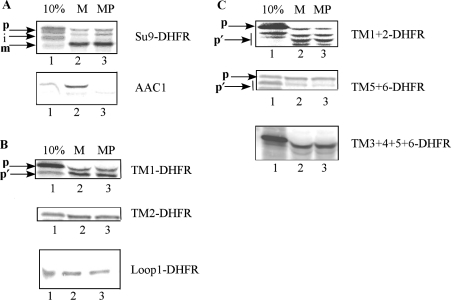
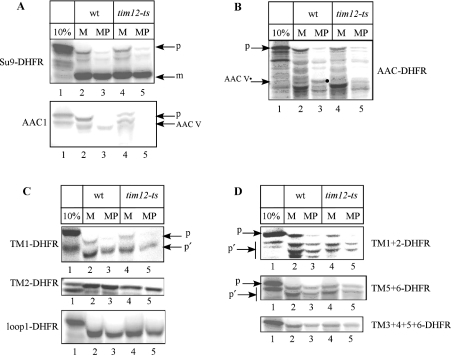
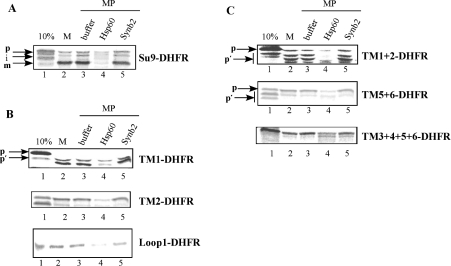
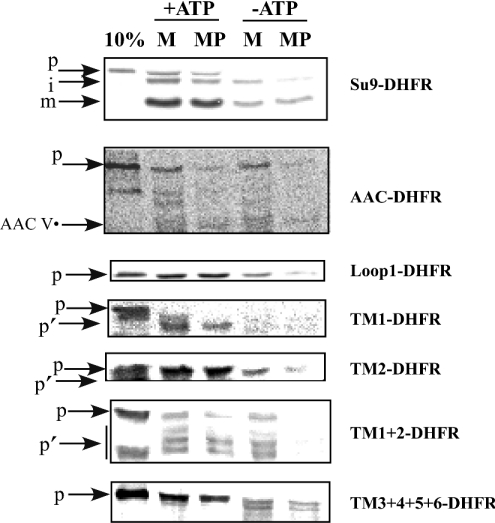
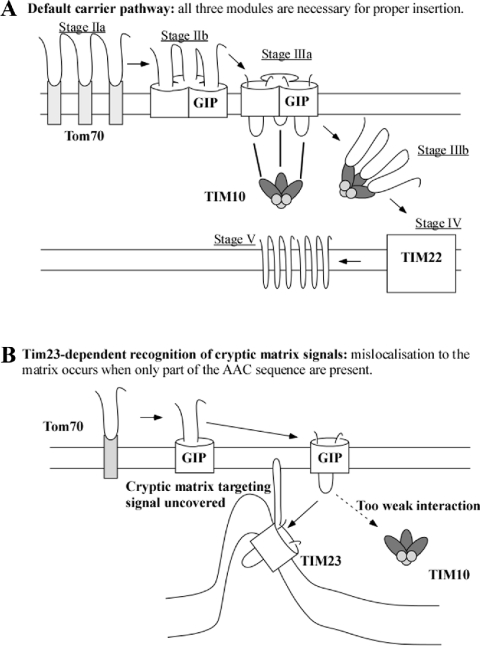
The yeast ADP/ATP carrier (AAC) is a mitochondrial protein that is targeted to the inner membrane via the TIM10 and TIM22 translocase complexes. AAC is devoid of a typical mitochondrial targeting signal and its targeting and insertion are thought to be guided by internal amino acid sequences. Here we show that AAC contains a cryptic matrix targeting signal that can target up to two thirds of the N-terminal part of the protein to the matrix. This event is coordinated by the TIM23 translocase and displays all the features of the matrix-targeting pathway. However, in the context of the whole protein, this signal is 'masked' and rendered non-functional as the polypeptide is targeted to the inner membrane via the TIM10 and TIM22 translocases. Our data suggest that after crossing the outer membrane the whole polypeptide chain of AAC is necessary to commit the precursor to the TIM22-mediated inner membrane insertion pathway.
Figures
Similar articles
-
Import of carrier proteins into the mitochondrial inner membrane mediated by Tim22.Nature. 1996 Dec 12;384(6609):582-5. doi: 10.1038/384582a0. Nature. 1996. PMID: 8955274
-
Mitochondrial import of the ADP/ATP carrier: the essential TIM complex of the intermembrane space is required for precursor release from the TOM complex.Mol Cell Biol. 2002 Nov;22(22):7780-9. doi: 10.1128/MCB.22.22.7780-7789.2002. Mol Cell Biol. 2002. PMID: 12391147 Free PMC article.
-
Transport of the ADP/ATP carrier of mitochondria from the TOM complex to the TIM22.54 complex.EMBO J. 1999 Jun 15;18(12):3214-21. doi: 10.1093/emboj/18.12.3214. EMBO J. 1999. PMID: 10369662 Free PMC article.
-
Protein import into and across the mitochondrial inner membrane: role of the TIM23 and TIM22 translocons.Biochim Biophys Acta. 2002 Sep 2;1592(1):25-34. doi: 10.1016/s0167-4889(02)00261-6. Biochim Biophys Acta. 2002. PMID: 12191765 Review.
-
Protein import into mitochondria.IUBMB Life. 2001 Sep-Nov;52(3-5):101-12. doi: 10.1080/15216540152845894. IUBMB Life. 2001. PMID: 11798021 Review.
Cited by
-
The mitochondrial carrier pathway transports non-canonical substrates with an odd number of transmembrane segments.BMC Biol. 2020 Jan 6;18(1):2. doi: 10.1186/s12915-019-0733-6. BMC Biol. 2020. PMID: 31907035 Free PMC article.
-
Folding and biogenesis of mitochondrial small Tim proteins.Int J Mol Sci. 2013 Aug 13;14(8):16685-705. doi: 10.3390/ijms140816685. Int J Mol Sci. 2013. PMID: 23945562 Free PMC article. Review.
-
A cryptic mitochondrial targeting motif in Atg4D links caspase cleavage with mitochondrial import and oxidative stress.Autophagy. 2012 Apr;8(4):664-76. doi: 10.4161/auto.19227. Epub 2012 Apr 1. Autophagy. 2012. PMID: 22441018 Free PMC article.
-
Biogenesis of Mitochondrial Metabolite Carriers.Biomolecules. 2020 Jul 7;10(7):1008. doi: 10.3390/biom10071008. Biomolecules. 2020. PMID: 32645990 Free PMC article. Review.
References
-
- Truscott K. N., Brandner K., Pfanner N. Mechanisms of protein import into mitochondria. Curr. Biol. 2003;13:R326–R337. - PubMed
-
- El Moualij B., Duyckaerts C., Lamotte-Brasseur J., Sluse F. E. Phylogenetic classification of the mitochondrial carrier family of Saccharomyces cerevisiae. Yeast. 1997;13:573–581. - PubMed
-
- Pfanner N., Neupert W. Distinct steps in the import of ADP/ATP carrier into mitochondria. J. Biol. Chem. 1987;262:7528–7536. - PubMed
-
- Zimmermann R., Neupert W. Transport of proteins into mitochondria. Posttranslational transfer of ADP/ATP carrier into mitochondria in vitro. Eur. J. Biochem. 1980;109:217–229. - PubMed
-
- Ryan M. T., Muller H., Pfanner N. Functional staging of ADP/ATP carrier translocation across the outer mitochondrial membrane. J. Biol. Chem. 1999;274:20619–20627. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases