

Loss of KCNJ10 protein expression abolishes endocochlear potential and causes deafness in Pendred syndrome mouse model
- PMID: 15320950
- PMCID: PMC516044
- DOI: 10.1186/1741-7015-2-30
Loss of KCNJ10 protein expression abolishes endocochlear potential and causes deafness in Pendred syndrome mouse model
Abstract
Background: Pendred syndrome, a common autosomal-recessive disorder characterized by congenital deafness and goiter, is caused by mutations of SLC26A4, which codes for pendrin. We investigated the relationship between pendrin and deafness using mice that have (Slc26a4+/+) or lack a complete Slc26a4 gene (Slc26a4-/-).
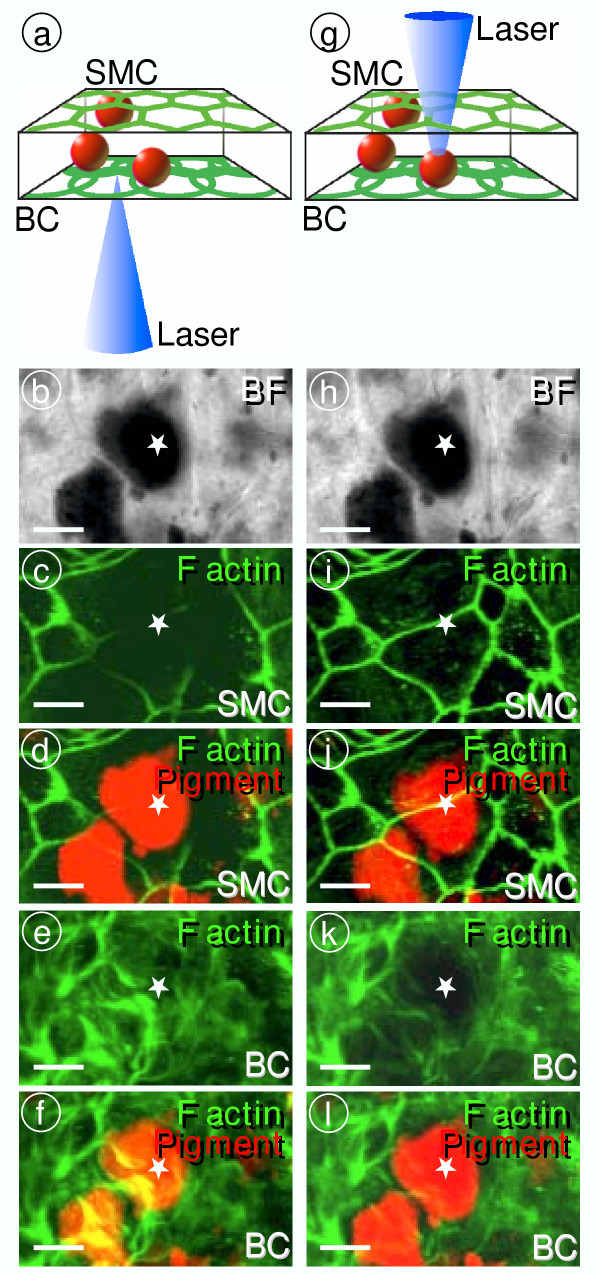
Methods: Expression of pendrin and other proteins was determined by confocal immunocytochemistry. Expression of mRNA was determined by quantitative RT-PCR. The endocochlear potential and the endolymphatic K+ concentration were measured with double-barreled microelectrodes. Currents generated by the stria marginal cells were recorded with a vibrating probe. Tissue masses were evaluated by morphometric distance measurements and pigmentation was quantified by densitometry.
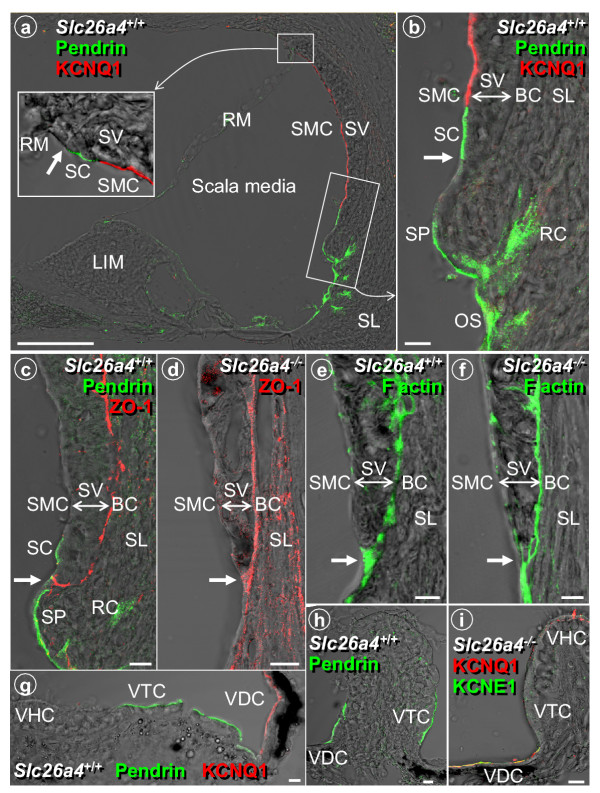
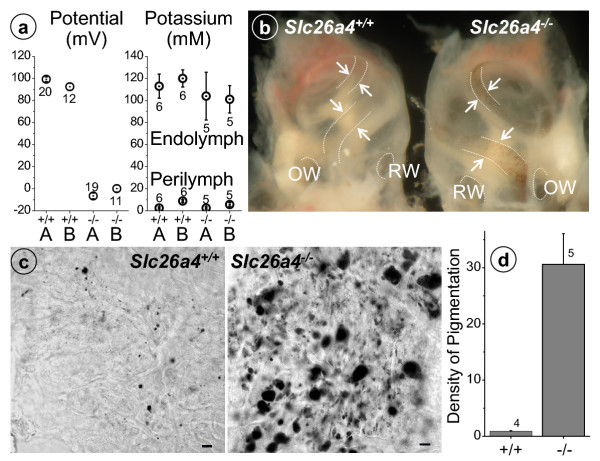
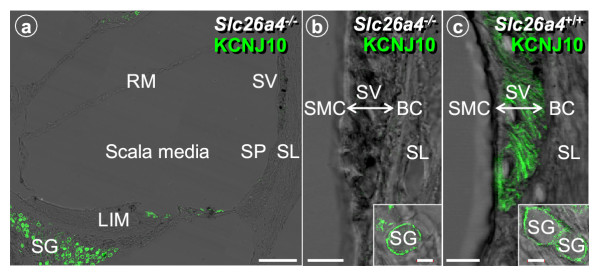
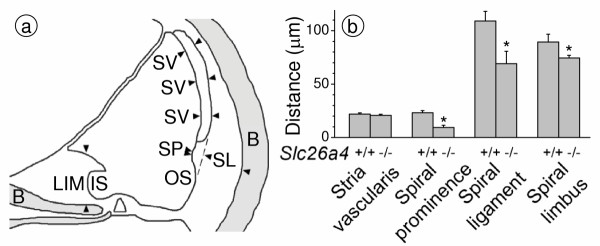
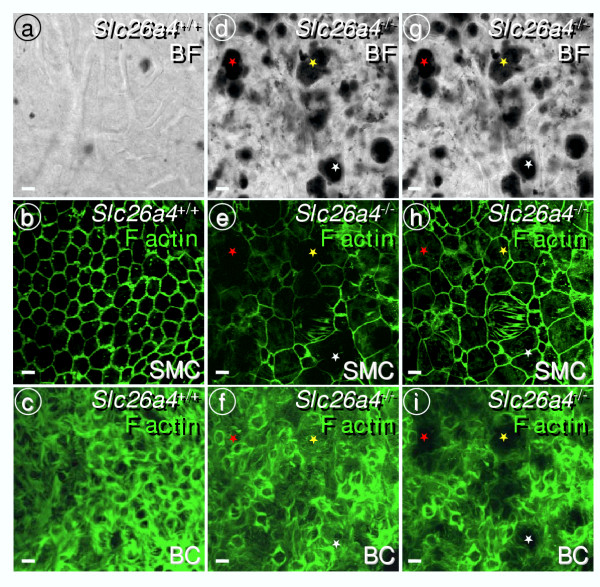
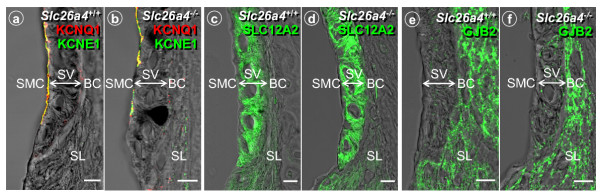
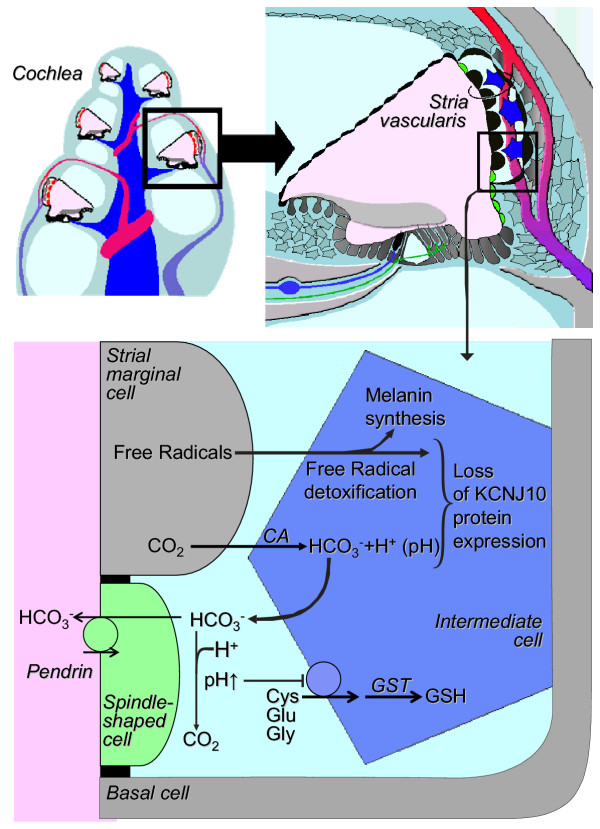
Results: Pendrin was found in the cochlea in apical membranes of spiral prominence cells and spindle-shaped cells of stria vascularis, in outer sulcus and root cells. Endolymph volume in Slc26a4-/- mice was increased and tissue masses in areas normally occupied by type I and II fibrocytes were reduced. Slc26a4-/- mice lacked the endocochlear potential, which is generated across the basal cell barrier by the K+ channel KCNJ10 localized in intermediate cells. Stria vascularis was hyperpigmented, suggesting unalleviated free radical damage. The basal cell barrier appeared intact; intermediate cells and KCNJ10 mRNA were present but KCNJ10 protein was absent. Endolymphatic K+ concentrations were normal and membrane proteins necessary for K+ secretion were present, including the K+ channel KCNQ1 and KCNE1, Na+/2Cl-/K+ cotransporter SLC12A2 and the gap junction GJB2.
Conclusions: These observations demonstrate that pendrin dysfunction leads to a loss of KCNJ10 protein expression and a loss of the endocochlear potential, which may be the direct cause of deafness in Pendred syndrome.
Figures

References
-
- Pendred V. Deaf-mutism and goitre. Lancet. 1896;11:532. doi: 10.1016/S0140-6736(01)74403-0. - DOI
-
- Royaux IE, Suzuki K, Mori A, Katoh R, Everett LA, Kohn LD, Green ED. Pendrin, the protein encoded by the Pendred syndrome gene (PDS), is an apical porter of iodide in the thyroid and is regulated by thyroglobulin in FRTL-5 cells. Endocrinology. 2000;141:839–845. doi: 10.1210/en.141.2.839. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
