

Contribution of Burkholderia cenocepacia flagella to infectivity and inflammation
- PMID: 15322006
- PMCID: PMC517433
- DOI: 10.1128/IAI.72.9.5126-5134.2004
Contribution of Burkholderia cenocepacia flagella to infectivity and inflammation
Abstract
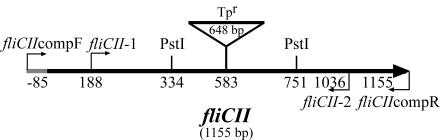
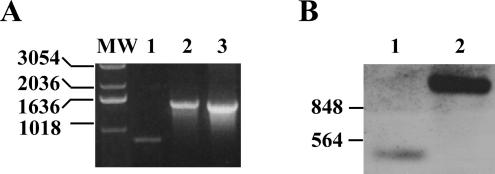
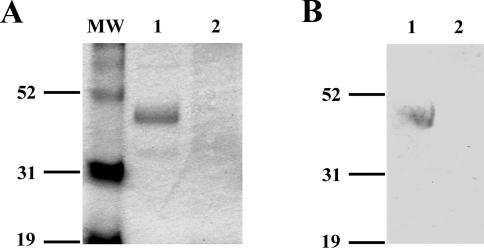
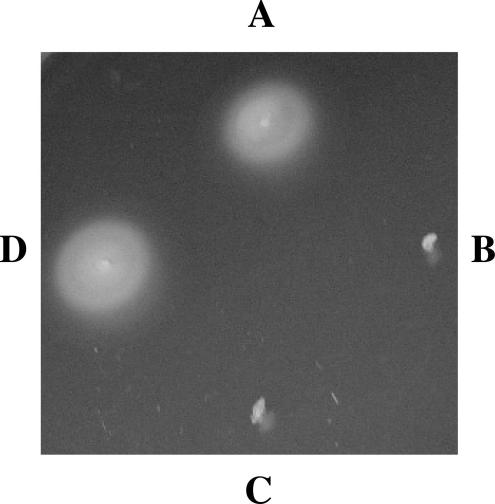
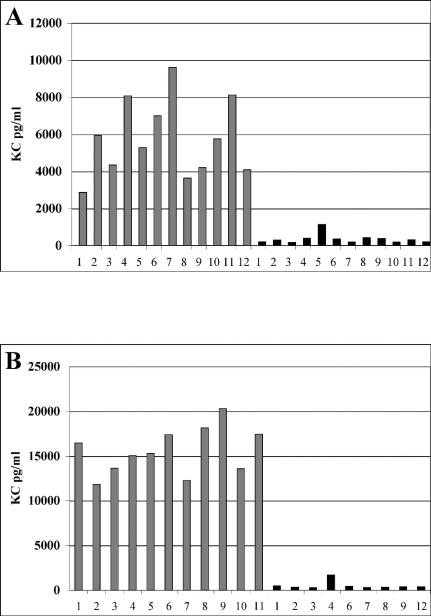
Burkholderia cenocepacia is an opportunistic pathogen that can cause severe lung infections in cystic fibrosis patients. To understand the contribution of B. cenocepacia flagella to infection, a strain mutated in the major flagellin subunit, fliCII, was constructed in B. cenocepacia K56-2 and tested in a murine agar bead model of lung infection. C57/BL6 mice infected with approximately 10(8) wild-type K56-2 bacteria exhibited 40% mortality after 3 days, whereas no mortality was noted in mice infected with the fliCII mutant. Among the mice surviving the infection with either strain, there was no significant difference in the bacterial loads in the lungs and spleen, bacteremia, weight loss, or infiltration of immune effector cells at 3 days postinfection. Similar results were observed at 24 h, prior to expression of the lethality phenotype. KC, a murine interleukin-8 (IL-8) homolog, was elevated in both the bronchoalveolar lavage fluid and serum of mice infected with the wild type compared to the fliCII mutant at 24 h, suggesting that flagella stimulated host cells. To demonstrate that flagella contributed to these responses, the interaction between B. cenocepacia and Toll-like receptor 5 (TLR5) was investigated. Infection of HEK293 cells with heat-killed wild-type K56-2, but not infection with the fliCII mutant, resulted in both NF-kappaB activation and IL-8 secretion that was dependent upon expression of TLR5. Together, these results demonstrate that B. cenocepacia flagella contribute to virulence in an in vivo infection model, and that induction of host immune responses through interaction with TLR5 may contribute to its overall pathogenic potential.
Figures



Similar articles
-
The CepIR quorum-sensing system contributes to the virulence of Burkholderia cenocepacia respiratory infections.Microbiology (Reading). 2003 Dec;149(Pt 12):3649-3658. doi: 10.1099/mic.0.26540-0. Microbiology (Reading). 2003. PMID: 14663096
-
Immune Recognition of the Epidemic Cystic Fibrosis Pathogen Burkholderia dolosa.Infect Immun. 2017 May 23;85(6):e00765-16. doi: 10.1128/IAI.00765-16. Print 2017 Jun. Infect Immun. 2017. PMID: 28348057 Free PMC article.
-
Lack of MyD88 protects the immunodeficient host against fatal lung inflammation triggered by the opportunistic bacteria Burkholderia cenocepacia.J Immunol. 2009 Jul 1;183(1):670-6. doi: 10.4049/jimmunol.0801497. Epub 2009 Jun 17. J Immunol. 2009. PMID: 19535624
-
Host evasion by Burkholderia cenocepacia.Front Cell Infect Microbiol. 2012 Jan 11;1:25. doi: 10.3389/fcimb.2011.00025. eCollection 2011. Front Cell Infect Microbiol. 2012. PMID: 22919590 Free PMC article. Review.
-
Burkholderia cenocepacia in cystic fibrosis: epidemiology and molecular mechanisms of virulence.Clin Microbiol Infect. 2010 Jul;16(7):821-30. doi: 10.1111/j.1469-0691.2010.03237.x. Clin Microbiol Infect. 2010. PMID: 20880411 Review.
Cited by
-
The role of toll-like receptors in acute and chronic lung inflammation.J Inflamm (Lond). 2010 Nov 25;7:57. doi: 10.1186/1476-9255-7-57. J Inflamm (Lond). 2010. PMID: 21108806 Free PMC article.
-
Transcriptional responses of Burkholderia cenocepacia to polymyxin B in isogenic strains with diverse polymyxin B resistance phenotypes.BMC Genomics. 2011 Sep 29;12:472. doi: 10.1186/1471-2164-12-472. BMC Genomics. 2011. PMID: 21955326 Free PMC article.
-
Swimming motility in a longitudinal collection of clinical isolates of Burkholderia cepacia complex bacteria from people with cystic fibrosis.PLoS One. 2014 Sep 9;9(9):e106428. doi: 10.1371/journal.pone.0106428. eCollection 2014. PLoS One. 2014. PMID: 25203161 Free PMC article.
-
The genome of Burkholderia cenocepacia J2315, an epidemic pathogen of cystic fibrosis patients.J Bacteriol. 2009 Jan;191(1):261-77. doi: 10.1128/JB.01230-08. Epub 2008 Oct 17. J Bacteriol. 2009. PMID: 18931103 Free PMC article.
-
A putative lateral flagella of the cystic fibrosis pathogen Burkholderia dolosa regulates swimming motility and host cytokine production.PLoS One. 2018 Jan 18;13(1):e0189810. doi: 10.1371/journal.pone.0189810. eCollection 2018. PLoS One. 2018. PMID: 29346379 Free PMC article.
References
-
- Baldwin, A. S., Jr. 1996. The NF-κB and IκB proteins: new discoveries and insights. Annu. Rev. Immunol. 14:649-683. - PubMed
-
- Barton, G. M., and R. Medzhitov. 2003. Toll-like receptor signaling pathways. Science 300:1524-1525. - PubMed
-
- Chiu, C. H., A. Ostry, and D. P. Speert. 2001. Invasion of murine respiratory epithelial cells in vivo by Burkholderia cepacia. J. Med. Microbiol. 50:594-601. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
