

Substrate-induced asymmetry and channel closure revealed by the apoenzyme structure of Mycobacterium tuberculosis phosphopantetheine adenylyltransferase
- PMID: 15322293
- PMCID: PMC2280004
- DOI: 10.1110/ps.04816904
Substrate-induced asymmetry and channel closure revealed by the apoenzyme structure of Mycobacterium tuberculosis phosphopantetheine adenylyltransferase
Abstract
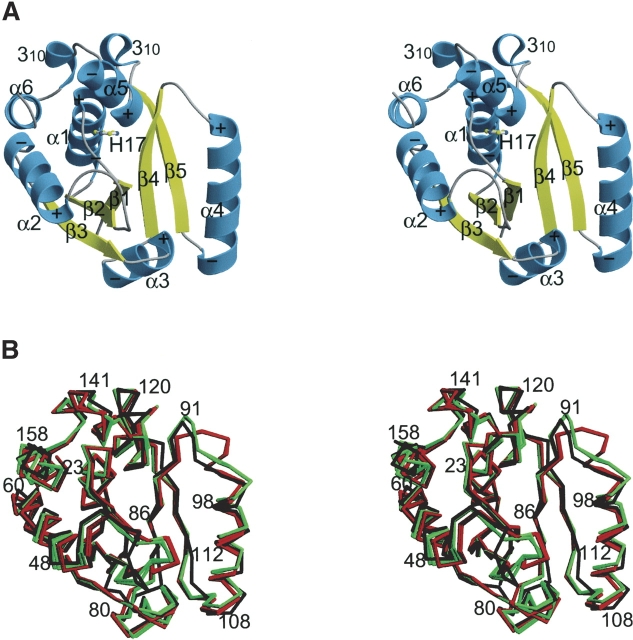
Phosphopantetheine adenylyltransferase (PPAT) catalyzes the penultimate step in prokaryotic coenzyme A (CoA) biosynthesis, directing the transfer of an adenylyl group from ATP to 4'-phosphopantetheine (Ppant) to yield dephospho-CoA (dPCoA). The crystal structures of Escherichia coli PPAT bound to its substrates, product, and inhibitor revealed an allosteric hexameric enzyme with half-of-sites reactivity, and established an in-line displacement catalytic mechanism. To provide insight into the mechanism of ligand binding we solved the apoenzyme (Apo) crystal structure of PPAT from Mycobacterium tuberculosis. In its Apo form, PPAT is a symmetric hexamer with an open solvent channel. However, ligand binding provokes asymmetry and alters the structure of the solvent channel, so that ligand binding becomes restricted to one trimer.
Figures



Similar articles
-
Structure of Mycobacterium tuberculosis phosphopantetheine adenylyltransferase in complex with the feedback inhibitor CoA reveals only one active-site conformation.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011 May 1;67(Pt 5):541-5. doi: 10.1107/S1744309111010761. Epub 2011 Apr 20. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011. PMID: 21543857 Free PMC article.
-
The crystal structure of a novel bacterial adenylyltransferase reveals half of sites reactivity.EMBO J. 1999 Apr 15;18(8):2021-30. doi: 10.1093/emboj/18.8.2021. EMBO J. 1999. PMID: 10205156 Free PMC article.
-
A novel adenylate binding site confers phosphopantetheine adenylyltransferase interactions with coenzyme A.J Bacteriol. 2003 Jul;185(14):4074-80. doi: 10.1128/JB.185.14.4074-4080.2003. J Bacteriol. 2003. PMID: 12837781 Free PMC article.
-
Phosphopantetheine Adenylyltransferase: A promising drug target to combat antibiotic resistance.Biochim Biophys Acta Proteins Proteom. 2021 Feb;1869(2):140566. doi: 10.1016/j.bbapap.2020.140566. Epub 2020 Nov 30. Biochim Biophys Acta Proteins Proteom. 2021. PMID: 33271445 Review.
-
The diverse world of coenzyme A binding proteins.Curr Opin Struct Biol. 1996 Dec;6(6):790-7. doi: 10.1016/s0959-440x(96)80009-1. Curr Opin Struct Biol. 1996. PMID: 8994879 Review.
Cited by
-
Experimentally validated novel inhibitors of Helicobacter pylori phosphopantetheine adenylyltransferase discovered by virtual high-throughput screening.PLoS One. 2013 Sep 5;8(9):e74271. doi: 10.1371/journal.pone.0074271. eCollection 2013. PLoS One. 2013. PMID: 24040220 Free PMC article.
-
Phosphopantetheine adenylyltransferase from Escherichia coli: investigation of the kinetic mechanism and role in regulation of coenzyme A biosynthesis.J Bacteriol. 2007 Nov;189(22):8196-205. doi: 10.1128/JB.00732-07. Epub 2007 Sep 14. J Bacteriol. 2007. PMID: 17873050 Free PMC article.
-
Structure of Mycobacterium tuberculosis phosphopantetheine adenylyltransferase in complex with the feedback inhibitor CoA reveals only one active-site conformation.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011 May 1;67(Pt 5):541-5. doi: 10.1107/S1744309111010761. Epub 2011 Apr 20. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011. PMID: 21543857 Free PMC article.
-
Biosynthesis of Pantothenic Acid and Coenzyme A.EcoSal Plus. 2007 Apr;2(2):10.1128/ecosalplus.3.6.3.4. doi: 10.1128/ecosalplus.3.6.3.4. EcoSal Plus. 2007. PMID: 26443589 Free PMC article.
-
The structure of Staphylococcus aureus phosphopantetheine adenylyltransferase in complex with 3'-phosphoadenosine 5'-phosphosulfate reveals a new ligand-binding mode.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009 Oct 1;65(Pt 10):987-91. doi: 10.1107/S1744309109036616. Epub 2009 Sep 23. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009. PMID: 19851003 Free PMC article.
References
-
- Begley, T.P., Kinsland, C., and Strauss, E. 2001. The biosynthesis of coenzyme A in bacteria. Vitam. Horm. 61 157–171. - PubMed
-
- Brown, K.L., Morris, V.K., and Izard, T. 2004. Rhombohedral crystals of Mycobacterium tuberculosis phosphopantetheine adenylyltransferase. Acta Crystallogr. D60 195–196. - PubMed
-
- Brünger, A.T., Adams, P.D., Clore, G.M., Delano, W.L., Gros, P., Grosse-Kunstleve, R.W., Jiang, J.-S., Kuszewski, J., Nilges, N., Pannu, N.S., et al. 1998. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D54 905–921. - PubMed
-
- Collaborative Computational Project, No. 4. 1994. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D50 760–763. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
