

Effects of Q/N-rich, polyQ, and non-polyQ amyloids on the de novo formation of the [PSI+] prion in yeast and aggregation of Sup35 in vitro
- PMID: 15326312
- PMCID: PMC516497
- DOI: 10.1073/pnas.0404968101
Effects of Q/N-rich, polyQ, and non-polyQ amyloids on the de novo formation of the [PSI+] prion in yeast and aggregation of Sup35 in vitro
Abstract
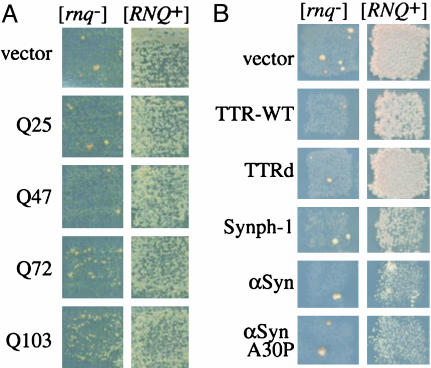
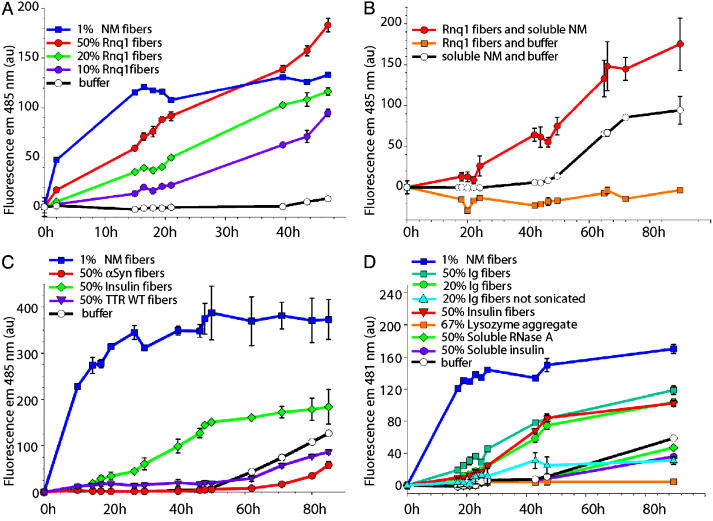
Prions are infectious protein conformations that are generally ordered protein aggregates. In the absence of prions, newly synthesized molecules of these same proteins usually maintain a conventional soluble conformation. However, prions occasionally arise even without a homologous prion template. The conformational switch that results in the de novo appearance of yeast prions with glutamine/aspargine (Q/N)-rich prion domains (e.g., [PSI+]), is promoted by heterologous prions with a similar domain (e.g., [RNQ+], also known as [PIN+]), or by overexpression of proteins with prion-like Q-, N-, or Q/N-rich domains. This finding led to the hypothesis that aggregates of heterologous proteins provide an imperfect template on which the new prion is seeded. Indeed, we show that newly forming Sup35 and preexisting Rnq1 aggregates always colocalize when [PSI+] appearance is facilitated by the [RNQ+] prion, and that Rnq1 fibers enhance the in vitro formation of fibers by the prion domain of Sup35 (NM). The proteins do not however form mixed, interdigitated aggregates. We also demonstrate that aggregating variants of the polyQ-containing domain of huntingtin promote the de novo conversion of Sup35 into [PSI+]; whereas nonaggregating variants of huntingtin and aggregates of non-polyQ amyloidogenic proteins, transthyretin, alpha-synuclein, and synphilin do not. Furthermore, transthyretin and alpha-synuclein amyloids do not facilitate NM aggregation in vitro, even though in [PSI+] cells NM and transthyretin aggregates also occasionally colocalize. Our data, especially the in vitro reproduction of the highly specific heterologous seeding effect, provide strong support for the hypothesis of cross-seeding in the spontaneous initiation of prion states.
Copyright 2004 The National Academy of Sciencs of the USA
Figures

Similar articles
-
Heterologous aggregates promote de novo prion appearance via more than one mechanism.PLoS Genet. 2015 Jan 8;11(1):e1004814. doi: 10.1371/journal.pgen.1004814. eCollection 2015 Jan. PLoS Genet. 2015. PMID: 25568955 Free PMC article.
-
Pathogenic polyglutamine tracts are potent inducers of spontaneous Sup35 and Rnq1 amyloidogenesis.PLoS One. 2010 Mar 10;5(3):e9642. doi: 10.1371/journal.pone.0009642. PLoS One. 2010. PMID: 20224794 Free PMC article.
-
Increased [PSI+] appearance by fusion of Rnq1 with the prion domain of Sup35 in Saccharomyces cerevisiae.Eukaryot Cell. 2009 Jul;8(7):968-76. doi: 10.1128/EC.00353-08. Epub 2009 May 1. Eukaryot Cell. 2009. PMID: 19411620 Free PMC article.
-
A bipolar personality of yeast prion proteins.Prion. 2011 Oct-Dec;5(4):305-10. doi: 10.4161/pri.18307. Epub 2011 Oct 1. Prion. 2011. PMID: 22156730 Free PMC article. Review.
-
Prion and nonprion amyloids: a comparison inspired by the yeast Sup35 protein.Prion. 2007 Jul-Sep;1(3):179-84. doi: 10.4161/pri.1.3.4840. Epub 2007 Jul 6. Prion. 2007. PMID: 19164899 Free PMC article. Review.
Cited by
-
The effects of amino acid composition on yeast prion formation and prion domain interactions.Prion. 2010 Apr-Jun;4(2):60-5. doi: 10.4161/pri.4.2.12190. Epub 2010 Apr 28. Prion. 2010. PMID: 20495349 Free PMC article.
-
Physiological and environmental control of yeast prions.FEMS Microbiol Rev. 2014 Mar;38(2):326-44. doi: 10.1111/1574-6976.12053. Epub 2013 Dec 4. FEMS Microbiol Rev. 2014. PMID: 24236638 Free PMC article. Review.
-
The self-interaction of native TDP-43 C terminus inhibits its degradation and contributes to early proteinopathies.Nat Commun. 2012 Apr 3;3:766. doi: 10.1038/ncomms1766. Nat Commun. 2012. PMID: 22473010
-
Aggregation and Prion-Inducing Properties of the G-Protein Gamma Subunit Ste18 are Regulated by Membrane Association.Int J Mol Sci. 2020 Jul 16;21(14):5038. doi: 10.3390/ijms21145038. Int J Mol Sci. 2020. PMID: 32708832 Free PMC article.
-
Structural Bases of Prion Variation in Yeast.Int J Mol Sci. 2022 May 20;23(10):5738. doi: 10.3390/ijms23105738. Int J Mol Sci. 2022. PMID: 35628548 Free PMC article. Review.
References
-
- Griffith, J. S. (1967) Nature 215, 1043–1044. - PubMed
-
- Prusiner, S. B. (1982) Science 216, 136–144. - PubMed
-
- Wickner, R. B. (1994) Science 264, 566–569. - PubMed
-
- Uptain, S. M. & Lindquist, S. (2002) Annu. Rev. Microbiol. 56, 703–741. - PubMed
-
- Serio, T. R. & Lindquist, S. L. (2001) Adv. Protein Chem. 59, 391–412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
