

Increased frequency of aberrant V(D)J recombination products in core RAG-expressing mice
- PMID: 15328366
- PMCID: PMC516053
- DOI: 10.1093/nar/gkh778
Increased frequency of aberrant V(D)J recombination products in core RAG-expressing mice
Abstract
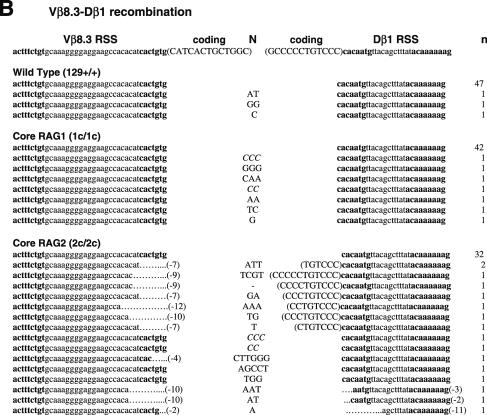
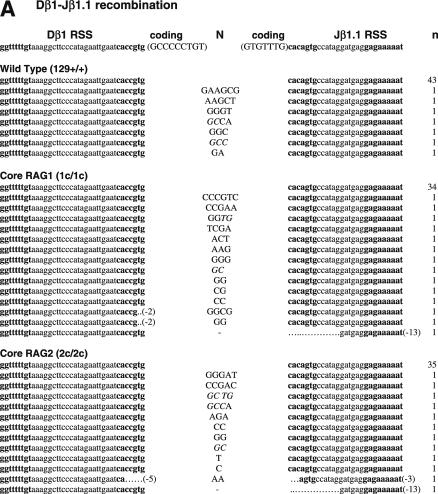
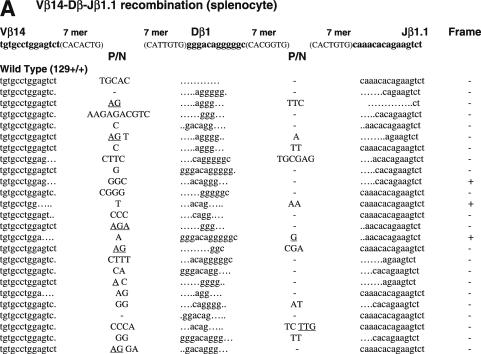
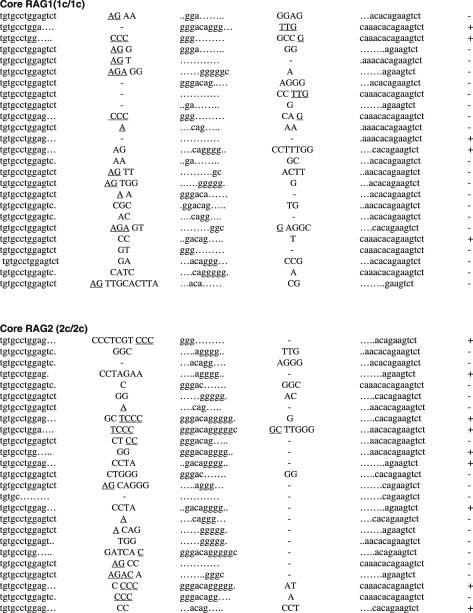
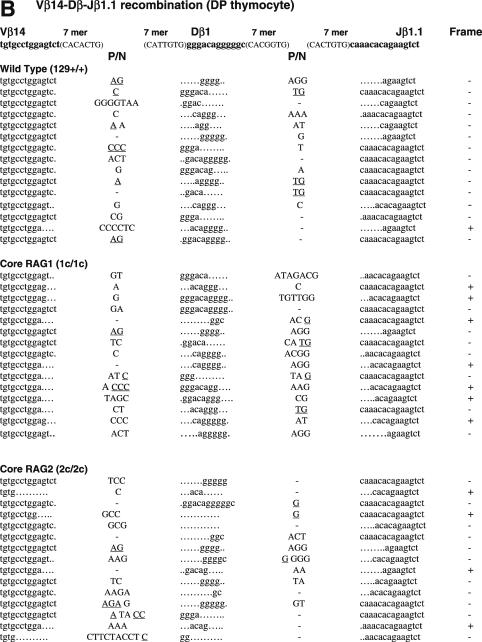
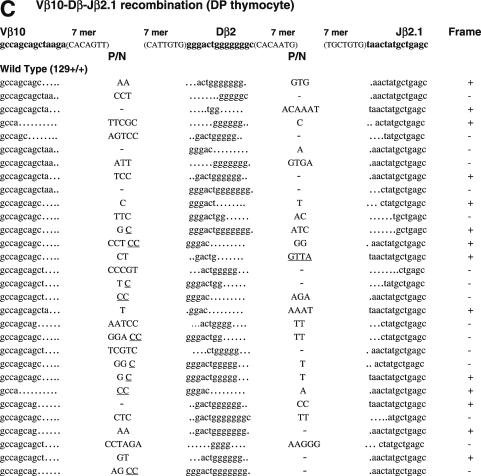
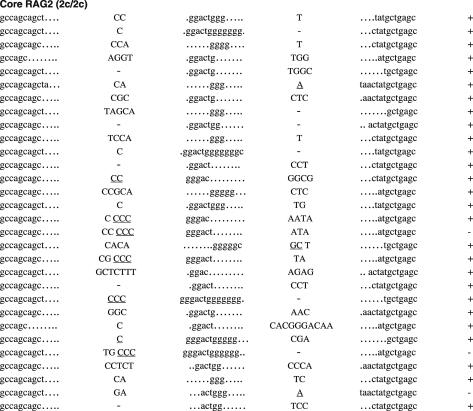
RAG1 and RAG2 play a central role in V(D)J recombination, a process for antigen receptor gene assembly. The truncated 'core' regions of RAGs are sufficient to catalyze the recombination reaction, although with lower joining efficiency than full-length proteins. To investigate the role of the non-core regions of RAGs in the end-joining phase of antigen receptor rearrangement, we analyzed recombination products isolated from core RAG1 and core RAG2 knock-in mice. Here, we report that the truncation of RAGs increases the frequency of aberrant recombination in vivo. Signal joints (SJs) associated with V-to-D recombination of core RAG1 knock-in mice were normal, whereas those of core RAG2 knock-in mice were highly imprecise, containing large deletions and additions, and in some cases coding sequences. In contrast, we found an elevated level of imprecise D-to-J associated SJs for both core RAG1- and RAG2-expressing mice. Likewise, sequences of coding joints (CJs) were also affected by the expression of core RAGs. Finally, sequences found at the junctions of rearranged T-cell receptor loci were highly influenced by differences in rearranging recombination signal sequence pairs. We provide the first evidence that the non-core regions of RAGs have critical functions in the proper assembly and resolution of recombination intermediates in endogenous antigen receptor loci.
Figures







References
-
- Bassing C.H., Swat,W. and Alt,F.W. (2002) The mechanism and regulation of chromosomal V(D)J recombination. Cell, 109 (Suppl.), S45–S55. - PubMed
-
- Gellert M. (2002) V(D)J recombination: RAG proteins, repair factors, and regulation. Annu. Rev. Biochem., 71, 101–132. - PubMed
-
- Agrawal A. and Schatz,D.G. (1997) RAG1 and RAG2 form a stable postcleavage synaptic complex with DNA containing signal ends in V(D)J recombination. Cell, 89, 43–53. - PubMed
-
- Hiom K. and Gellert,M. (1998) Assembly of a 12/23 paired signal complex: a critical control point in V(D)J recombination. Mol. Cell, 1, 1011–1019. - PubMed
