

Chromosome healing through terminal deletions generated by de novo telomere additions in Saccharomyces cerevisiae
- PMID: 15328403
- PMCID: PMC516557
- DOI: 10.1073/pnas.0405443101
Chromosome healing through terminal deletions generated by de novo telomere additions in Saccharomyces cerevisiae
Abstract
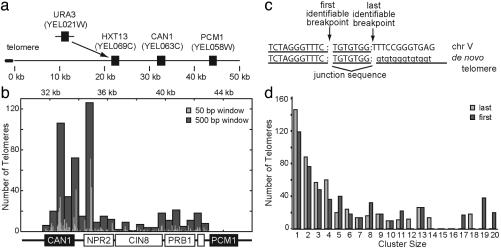
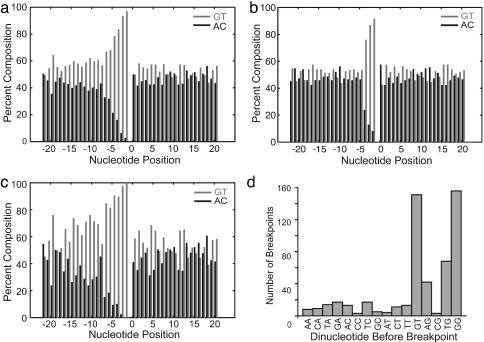
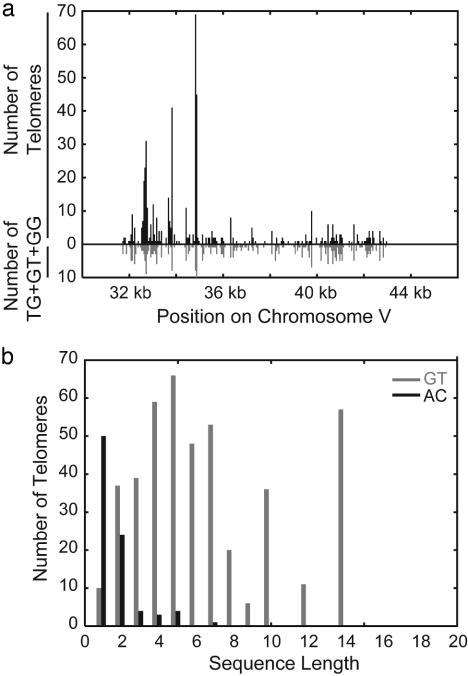
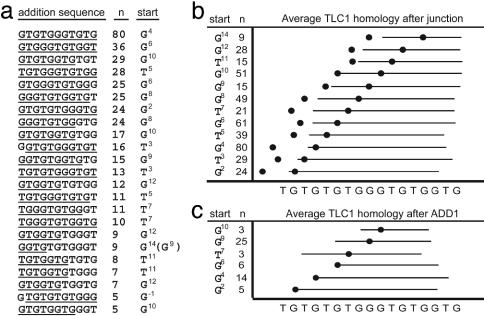
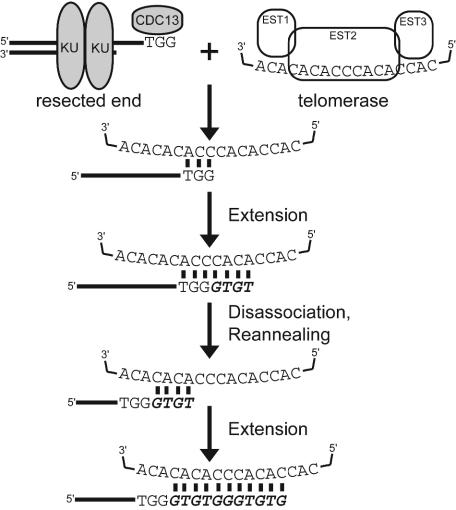
Broken chromosomes healed by de novo addition of a telomere are a major class of genome rearrangements seen in Saccharomyces cerevisiae and similar to rearrangements seen in human tumors. We have analyzed the sequences of 534 independent de novo telomere additions within a 12-kb region of chromosome V. The distribution of events mirrored that of four-base sequences consisting of the GG, GT, and TG dinucleotides, suggesting that de novo telomere additions occur at short regions of homology to the telomerase guide RNA. These chromosomal sequences restrict potential registrations of the added telomere sequence. The first 11 nucleotides of the addition sequences fell into common families that included 91% of the breakpoints. The observed registrations suggest that the 3' end of the TLC1 guide RNA is involved in annealing but not as a template for synthesis. Some families of added sequences can be accounted for by one cycle of annealing and extension, whereas others require a minimum of two. The same pattern emerges for sequences added onto the most common addition sequence, indicating that de novo telomeres are added and extended by the same process. Together, these data indicate that annealing is central to telomerase registration, which limits telomere heterogeneity and resolves the problem of synthesizing Rap1 binding sites by a nonprocessive telomerase with a low-complexity guide RNA sequence.
Figures

Similar articles
-
Chromosome healing by de novo telomere addition in Saccharomyces cerevisiae.Mol Microbiol. 2006 Mar;59(5):1357-68. doi: 10.1111/j.1365-2958.2006.05026.x. Mol Microbiol. 2006. PMID: 16468981 Review.
-
Analysis of De Novo Telomere Addition by Southern Blot.Methods Mol Biol. 2018;1672:363-373. doi: 10.1007/978-1-4939-7306-4_25. Methods Mol Biol. 2018. PMID: 29043636
-
De novo synthesis of telomere sequences at the healed breakpoints of wheat deletion chromosomes.Mol Gen Genet. 1999 Dec;262(4-5):851-6. doi: 10.1007/s004380051150. Mol Gen Genet. 1999. PMID: 10628870
-
When the Ends Justify the Means: Regulation of Telomere Addition at Double-Strand Breaks in Yeast.Front Cell Dev Biol. 2021 Mar 18;9:655377. doi: 10.3389/fcell.2021.655377. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33816507 Free PMC article. Review.
-
Characterization of terminal deletions at 7q32 and 22q13.3 healed by De novo telomere addition.Am J Hum Genet. 2000 Sep;67(3):610-22. doi: 10.1086/303050. Epub 2000 Aug 1. Am J Hum Genet. 2000. PMID: 10924407 Free PMC article.
Cited by
-
The Saccharomyces cerevisiae Rad6 postreplication repair and Siz1/Srs2 homologous recombination-inhibiting pathways process DNA damage that arises in asf1 mutants.Mol Cell Biol. 2009 Oct;29(19):5226-37. doi: 10.1128/MCB.00894-09. Epub 2009 Jul 27. Mol Cell Biol. 2009. PMID: 19635810 Free PMC article.
-
Defective resection at DNA double-strand breaks leads to de novo telomere formation and enhances gene targeting.PLoS Genet. 2010 May 13;6(5):e1000948. doi: 10.1371/journal.pgen.1000948. PLoS Genet. 2010. PMID: 20485519 Free PMC article.
-
Stimulation of gross chromosomal rearrangements by the human CEB1 and CEB25 minisatellites in Saccharomyces cerevisiae depends on G-quadruplexes or Cdc13.PLoS Genet. 2012;8(11):e1003033. doi: 10.1371/journal.pgen.1003033. Epub 2012 Nov 1. PLoS Genet. 2012. PMID: 23133402 Free PMC article.
-
A genetic and structural study of genome rearrangements mediated by high copy repeat Ty1 elements.PLoS Genet. 2011 May;7(5):e1002089. doi: 10.1371/journal.pgen.1002089. Epub 2011 May 26. PLoS Genet. 2011. PMID: 21637792 Free PMC article.
-
DNA repair pathway selection caused by defects in TEL1, SAE2, and de novo telomere addition generates specific chromosomal rearrangement signatures.PLoS Genet. 2014 Apr 3;10(4):e1004277. doi: 10.1371/journal.pgen.1004277. eCollection 2014 Apr. PLoS Genet. 2014. PMID: 24699249 Free PMC article.
References
-
- Mitelman, F. (1991) Catalog of Chromosome Aberrations in Cancer (Wiley-Liss, New York).
-
- Kolodner, R. & Marsischky, G. (1999) Curr. Opin. Genet. Dev. 9, 89-96. - PubMed
-
- Thompson, L. & Schild, D. (2002) Mutat. Res. 509, 49-78. - PubMed
-
- Mohaghegh, P. & Hickson, I. (2002) Int. J. Biochem. Cell Biol. 34, 1496-1501. - PubMed
-
- Friedberg, E. (2001) Nat. Rev. Cancer 1, 22-33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
