

Interdependency of brassinosteroid and auxin signaling in Arabidopsis
- PMID: 15328536
- PMCID: PMC509407
- DOI: 10.1371/journal.pbio.0020258
Interdependency of brassinosteroid and auxin signaling in Arabidopsis
Abstract
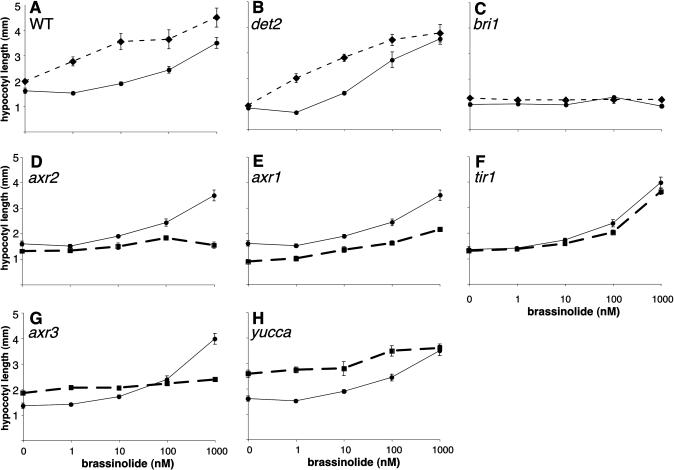
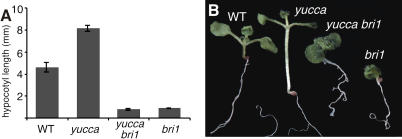
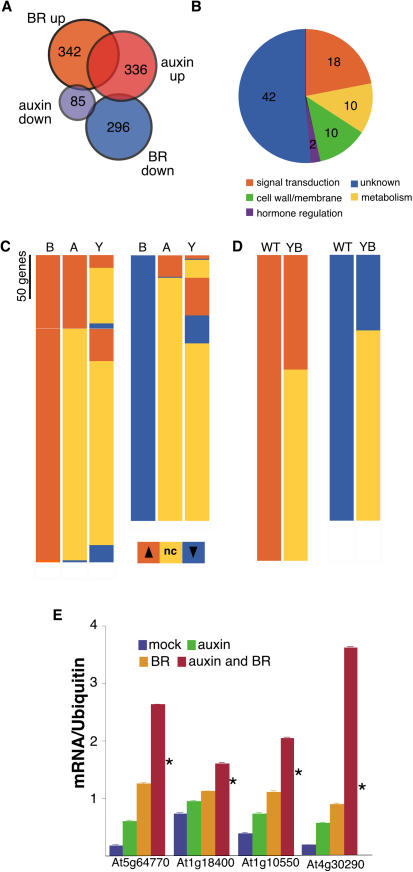
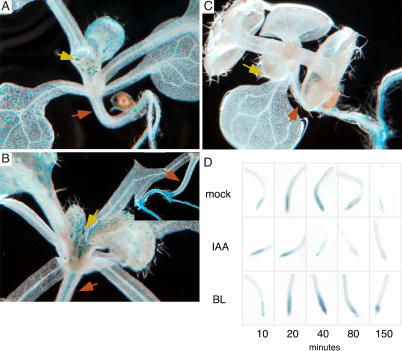
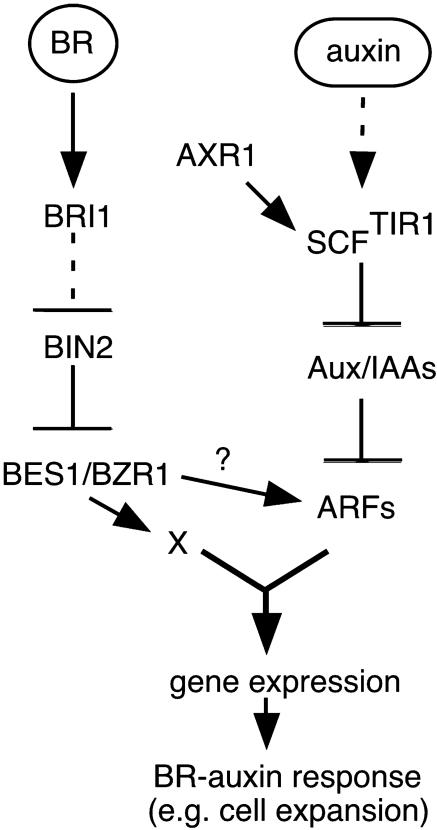
How growth regulators provoke context-specific signals is a fundamental question in developmental biology. In plants, both auxin and brassinosteroids (BRs) promote cell expansion, and it was thought that they activated this process through independent mechanisms. In this work, we describe a shared auxin:BR pathway required for seedling growth. Genetic, physiological, and genomic analyses demonstrate that response from one pathway requires the function of the other, and that this interdependence does not act at the level of hormone biosynthetic control. Increased auxin levels saturate the BR-stimulated growth response and greatly reduce BR effects on gene expression. Integration of these two pathways is downstream from BES1 and Aux/IAA proteins, the last known regulatory factors acting downstream of each hormone, and is likely to occur directly on the promoters of auxin:BR target genes. We have developed a new approach to identify potential regulatory elements acting in each hormone pathway, as well as in the shared auxin:BR pathway. We show that one element highly overrepresented in the promoters of auxin- and BR-induced genes is responsive to both hormones and requires BR biosynthesis for normal expression. This work fundamentally alters our view of BR and auxin signaling and describes a powerful new approach to identify regulatory elements required for response to specific stimuli.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
References
-
- Abel S, Nguyen MD, Theologis A. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana . J Mol Biol. 1995;251:533–549. - PubMed
-
- Alonso JM, Ecker JR. Sci STKE 2001: RE1; 2001. The ethylene pathway: A paradigm for plant hormone signaling and interaction. - PubMed
-
- Bailey TL, Elkan C. The value of prior knowledge in discovering motifs with MEME. Proc Int Conf Intell Syst Mol Biol. 1995;3:21–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
