

Epidermal growth factor and interleukin-1beta utilize divergent signaling pathways to synergistically upregulate cyclooxygenase-2 gene expression in human amnion-derived WISH cells
- PMID: 15329330
- PMCID: PMC1389598
- DOI: 10.1095/biolreprod.104.030841
Epidermal growth factor and interleukin-1beta utilize divergent signaling pathways to synergistically upregulate cyclooxygenase-2 gene expression in human amnion-derived WISH cells
Abstract
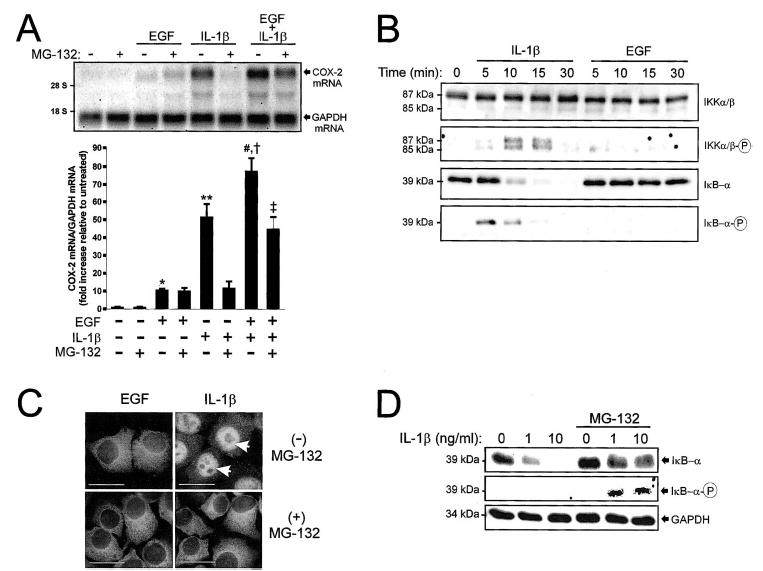
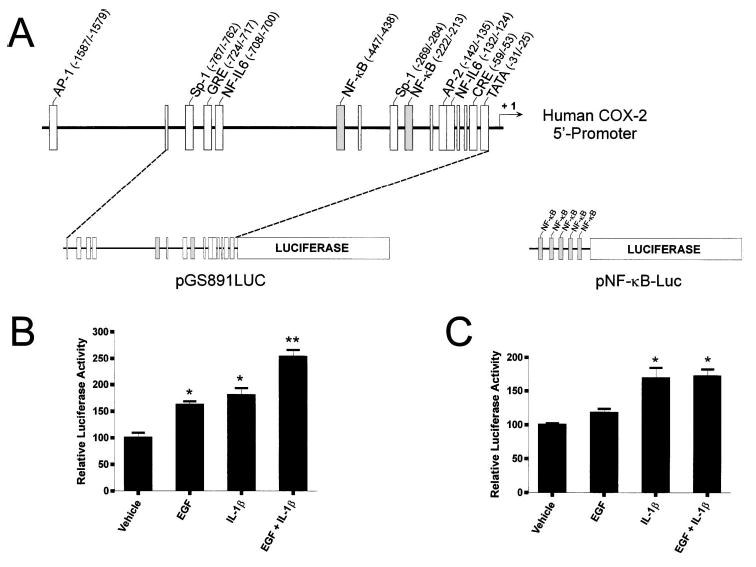
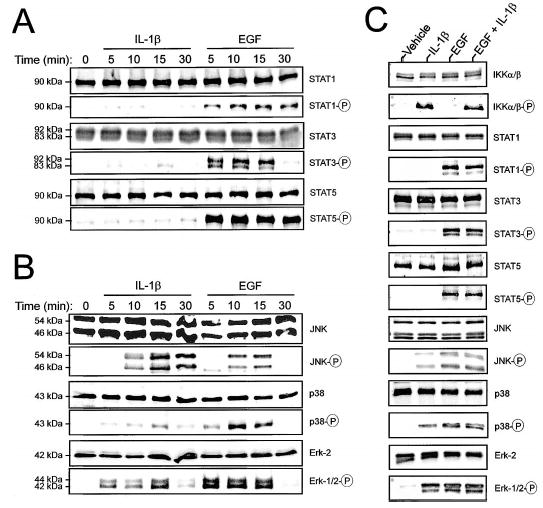
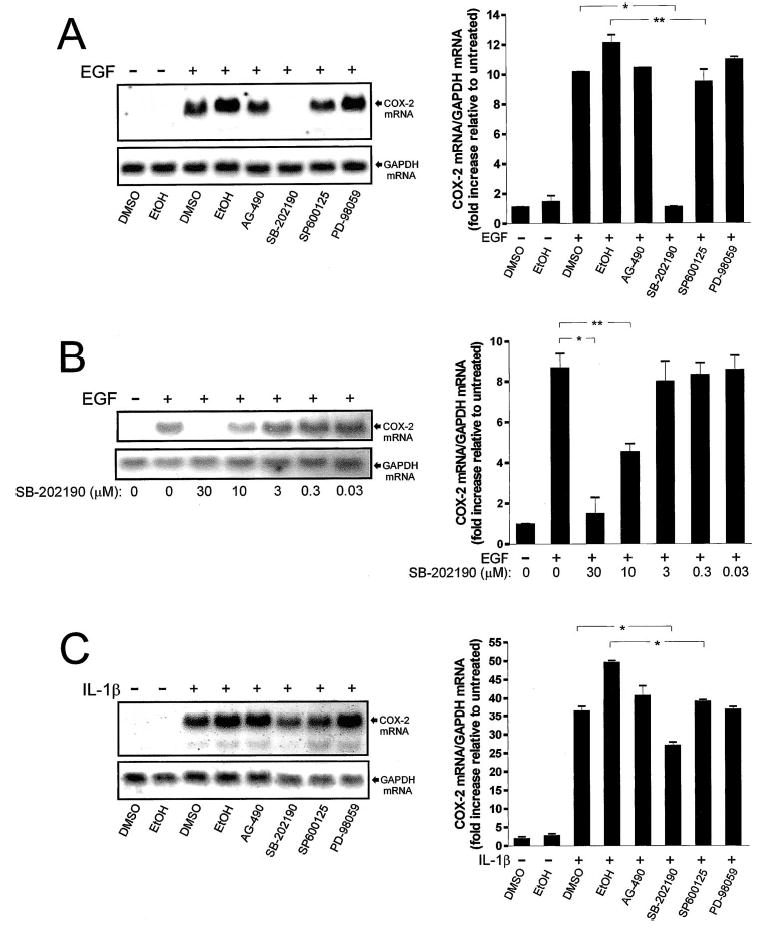
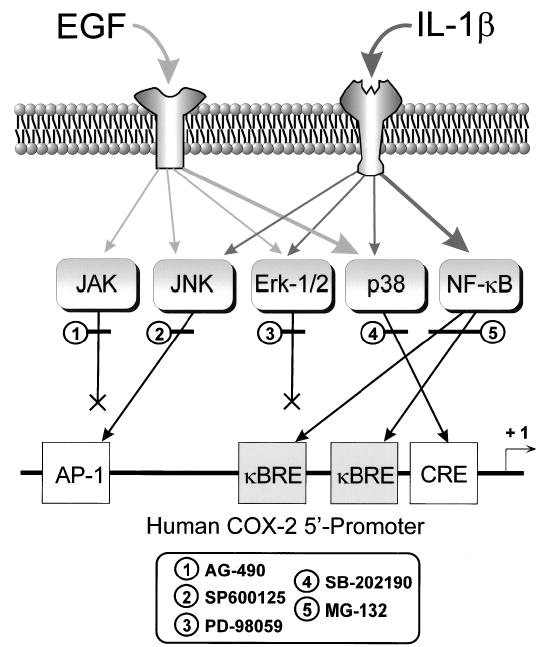
In human parturition, uterotonic prostaglandins (PGs) arise predominantly via increased expression of cyclooxygenase-2 (COX-2 [also known as prostaglandin synthase 2]) within intrauterine tissues. Interleukin-1 (IL-1) and epidermal growth factor (EGF), both inducers of COX-2 transcription, are among numerous factors that accumulate within amniotic fluid with advancing gestation. It was previously demonstrated that EGF could potentiate IL-1beta-driven PGE(2) production in amnion and amnion-derived (WISH) cells. To define the mechanism for this observation, we hypothesized that EGF and IL-1beta might exhibit synergism in regulating COX-2 gene expression. In WISH cells, combined treatment with EGF and IL-1beta resulted in a greater-than-additive increase in COX-2 mRNA relative to challenge with either agent independently. Augmentation of IL-1beta-induced transactivation by EGF was not observed in cells harboring reporter plasmids bearing nuclear factor-kappa B (NFkappaB) regulatory elements alone, but was evident when a fragment (-891/ +9) of the COX-2 gene 5'-promoter was present. Both agents transiently activated intermediates of multiple signaling pathways potentially involved in the regulation of COX-2 gene expression. The 26 S proteasome inhibitor, MG-132, selectively abrogated IL-1beta-driven NFkappaB activation and COX-2 mRNA expression. Only pharmacologic blockade of the p38 mitogen-activated protein kinase eliminated COX-2 expression following EGF stimulation. We conclude that EGF and IL-1beta appear to signal through different signaling cascades leading to COX-2 gene expression. IL-1beta employs the NFkappaB pathway predominantly, while the spectrum of EGF signaling is broader and includes p38 kinase. The synergism observed between IL-1beta and EGF does not rely on augmented NFkappaB function, but rather, occurs through differential use of independent response elements within the COX-2 promoter.
Figures
References
-
- Kniss DA, Iams JD. Regulation of parturition update. Endocrine and paracrine effectors of term and preterm labor. Clin Perinatol. 1998;25:819–836. - PubMed
-
- Kniss DA. Cyclooxygenases in reproductive medicine and biology. J Soc Gynecol Invest. 1999;6:285–292. - PubMed
-
- Johnson RF, Mitchell CM, Giles WB, Walters WA, Zakar T. The in vivo control of prostaglandin H synthase-2 messenger ribonucleic acid expression in the human amnion at parturition. J Clin Endocrinol Metab. 2002;87:2816–2823. - PubMed
-
- Keelan JA, Blumenstein M, Helliwell RJ, Sato TA, Marvin KW, Mitchell MD. Cytokines, prostaglandins and parturition—a review. Placenta. 2003;24(suppl A):S33–S46. - PubMed
-
- Bowen JM, Chamley L, Keelan JA, Mitchell MD. Cytokines of the placenta and extra-placental membranes: roles and regulation during human pregnancy and parturition. Placenta. 2002;23:257–273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
