

Activation and habituation of extracellular signal-regulated kinase phosphorylation in zebra finch auditory forebrain during song presentation
- PMID: 15329397
- PMCID: PMC6729643
- DOI: 10.1523/JNEUROSCI.1405-04.2004
Activation and habituation of extracellular signal-regulated kinase phosphorylation in zebra finch auditory forebrain during song presentation
Abstract
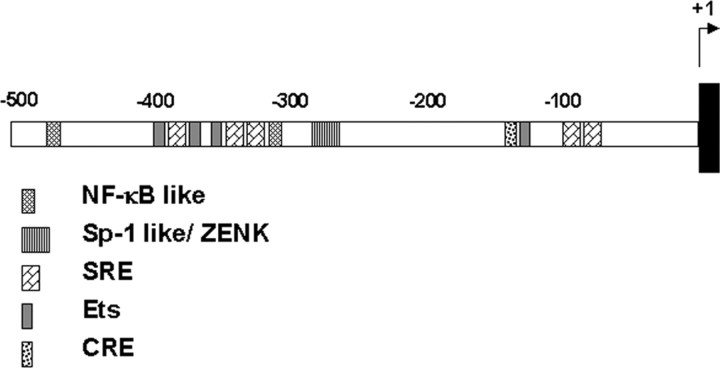
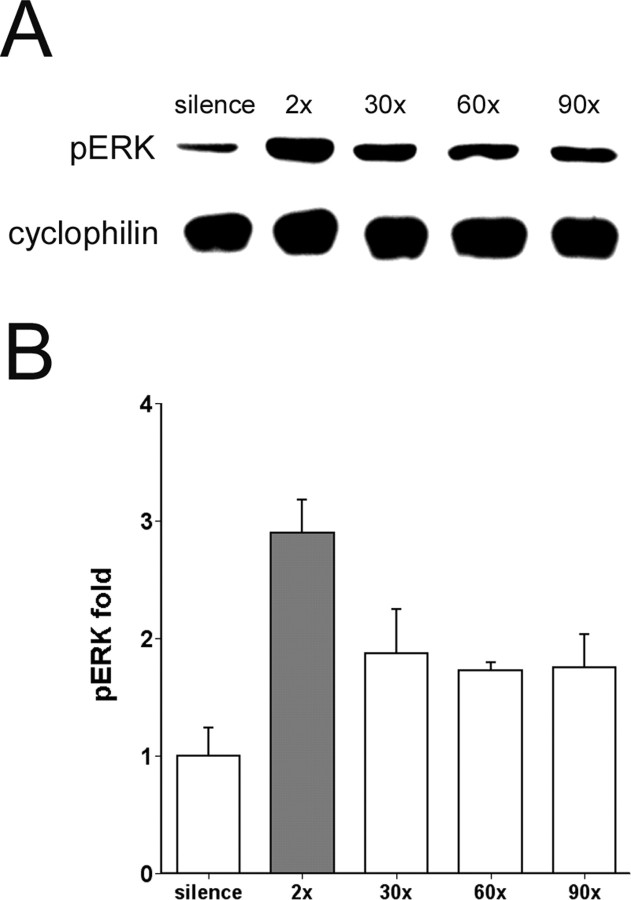
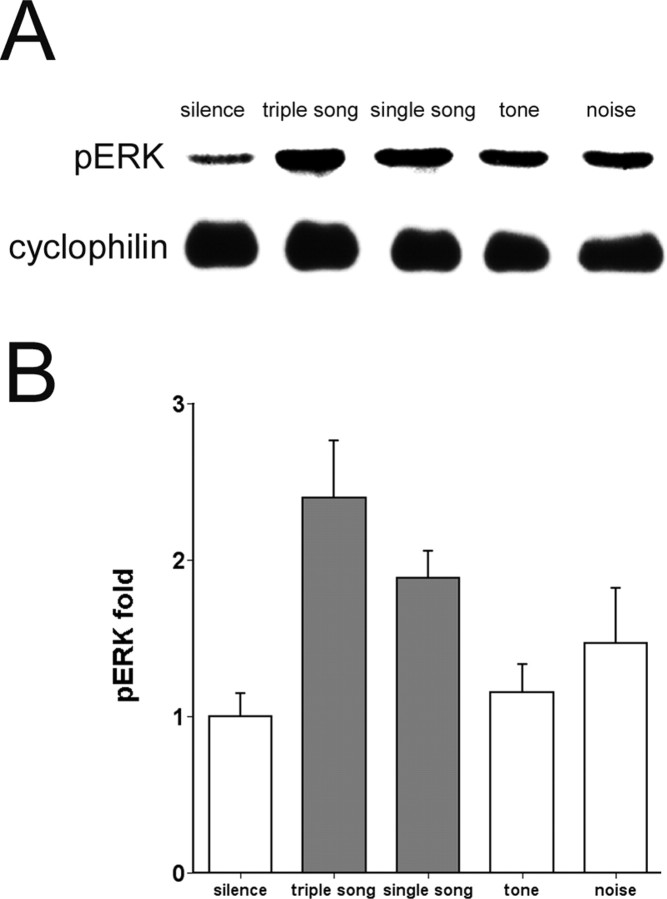
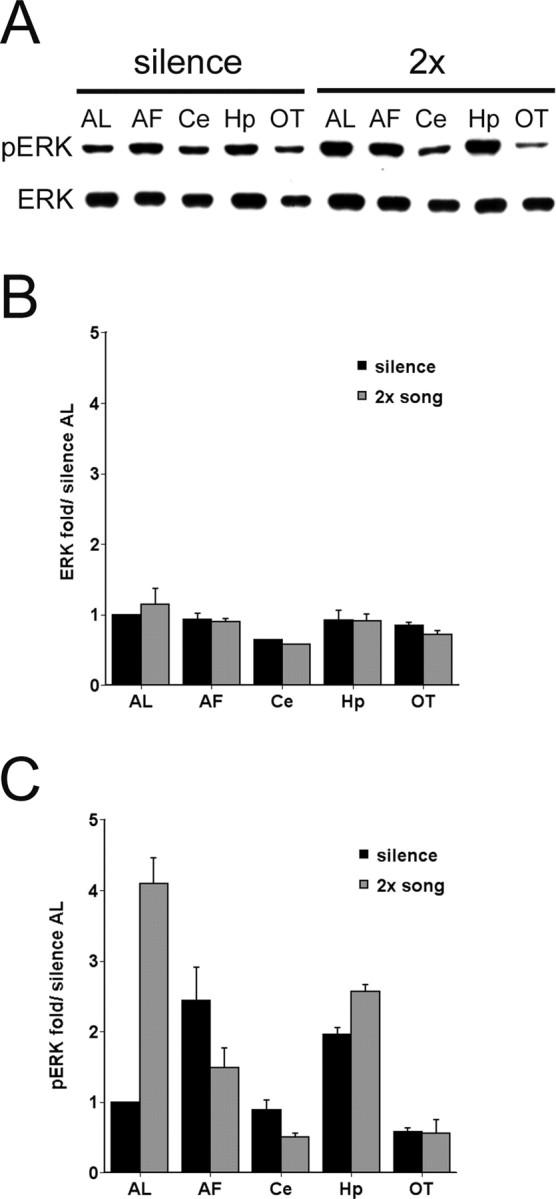
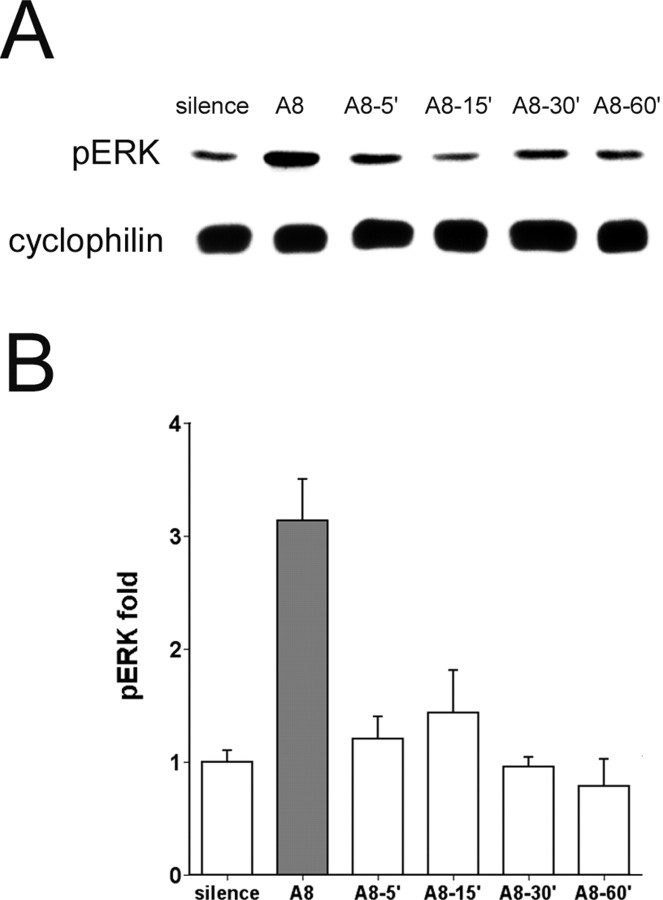
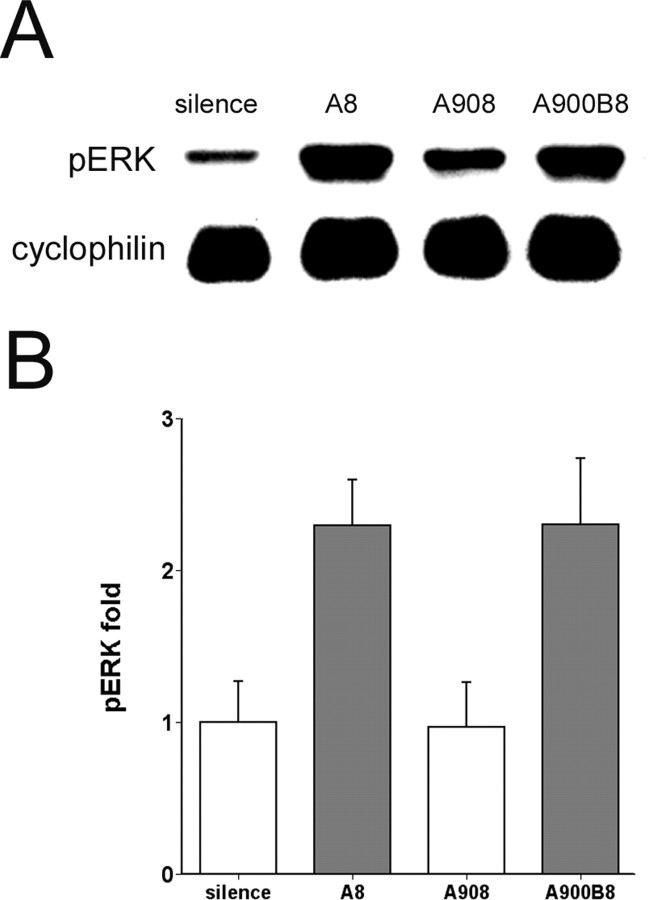
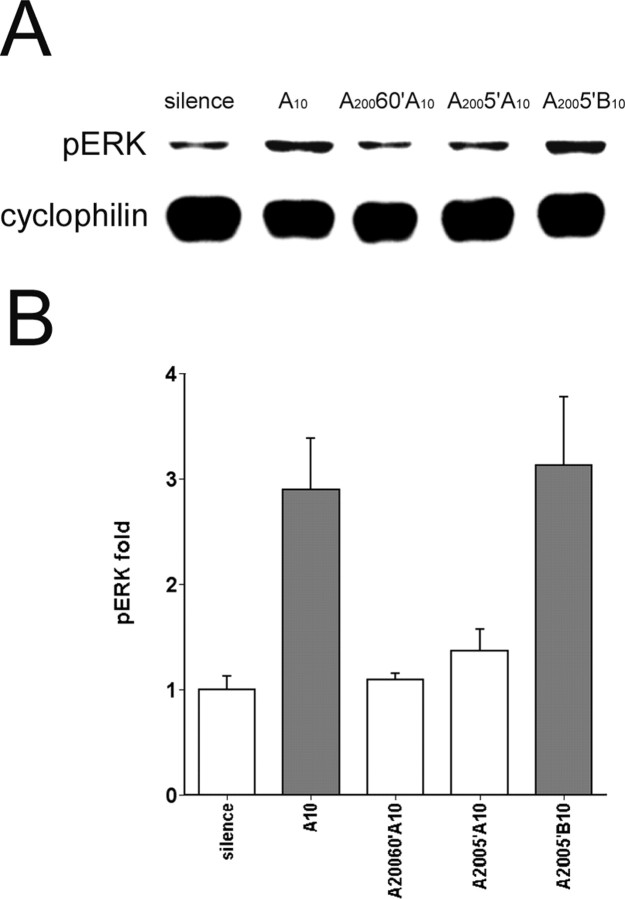
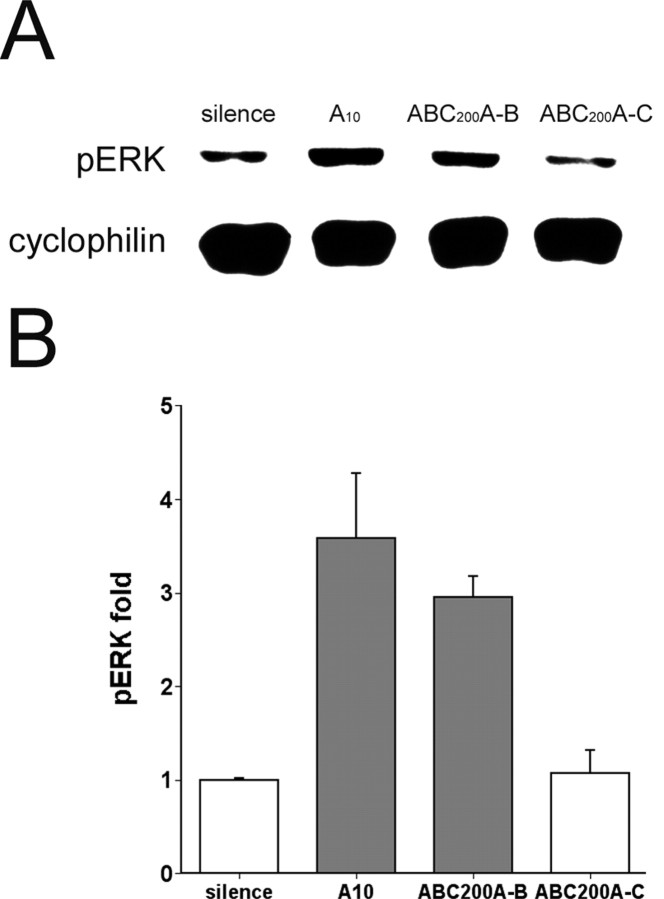
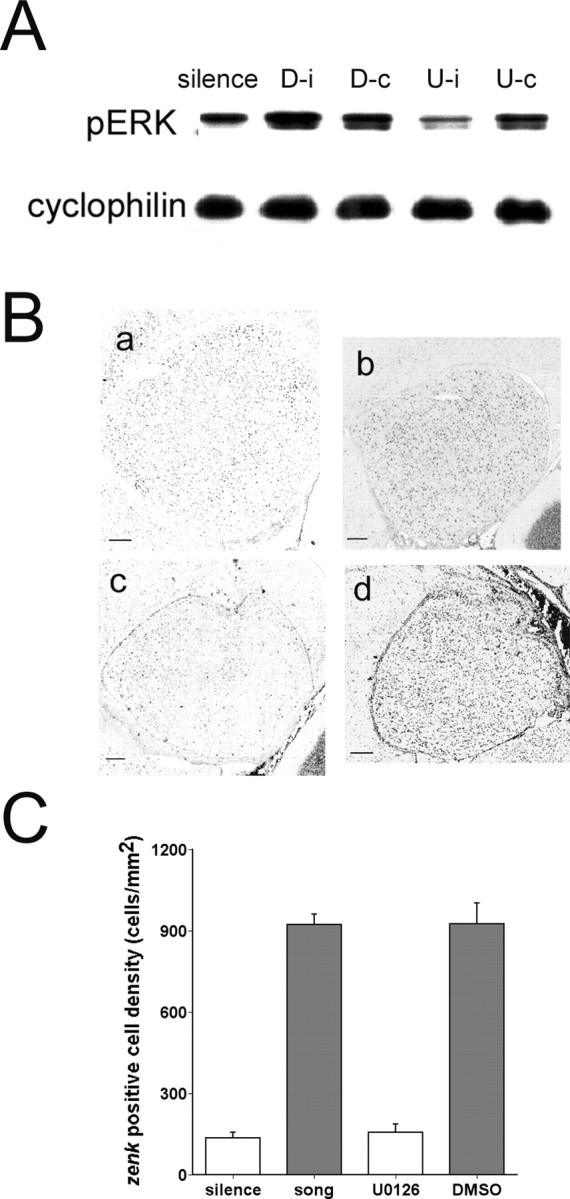
The sound of tape-recorded birdsong triggers a set of behavioral and physiological responses in zebra finches, including transcriptional activation of the zenk gene in the auditory forebrain. Song repetition leads to the stimulus-specific habituation of these responses. To gain insight into the mechanisms that couple auditory experience to gene regulation, we monitored the phosphorylation of the zebra finch extracellular signal-regulated kinase (ERK) protein by immunoblotting. Initial presentations of novel song (but not tones or noise) resulted in a rapid increase in ERK phosphorylation, followed by a return to basal levels within 5 min. This response was localized to the auditory forebrain where the zenk gene is activated. Sustained repetition of one song caused a selective habituation of the ERK response: a different song triggered another cycle of ERK phosphorylation without altering the habituated response to the first. To test directly for a role of ERK in experience-dependent zenk gene regulation, we infused an inhibitor of mitogen-activated and extracellular-regulated protein kinase kinase (MEK-1; the enzyme responsible for ERK activation) unilaterally into one auditory lobule just before song stimulation. The song-induced increase in zenk mRNA was blocked on the side of the injection, but not on the contralateral (uninfused) side. These results show that ERK phosphorylation is necessary for the initiation of the zenk gene response to novel song and identify ERK as a plausible site of signal integration underlying the selective habituation of genomic responses to a repeated song.
Figures
References
-
- Adams JP, Sweatt JD (2002) Molecular psychology: roles for the ERK MAP kinase cascade in memory. Ann Rev Pharmacol Toxicol 42: 135-163. - PubMed
-
- Alonso M, Viola H, Izquierdo I, Medina JH (2002) Aversive experiences are associated with a rapid and transient activation of ERKs in the rat hippocampus. Neurobiol Learn Mem 77: 119-124. - PubMed
-
- Athos J, Impey S, Pineda VV, Chen X, Storm DR (2002) Hippocampal CRE-mediated gene expression is required for contextual memory formation. Nat Neurosci 5: 1119-1120. - PubMed
-
- Atkins CM, Selcher JC, Petraitis JJ, Trzaskos JM, Sweatt JD (1998) The MAPK cascade is required for mammalian associative learning. Nat Neurosci 1: 602-609. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous