

Structural basis of actin sequestration by thymosin-beta4: implications for WH2 proteins
- PMID: 15329672
- PMCID: PMC517612
- DOI: 10.1038/sj.emboj.7600372
Structural basis of actin sequestration by thymosin-beta4: implications for WH2 proteins
Abstract
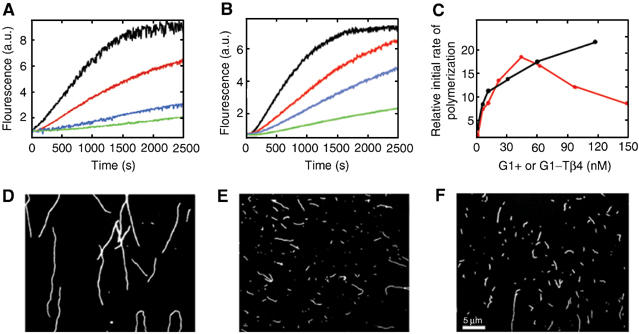
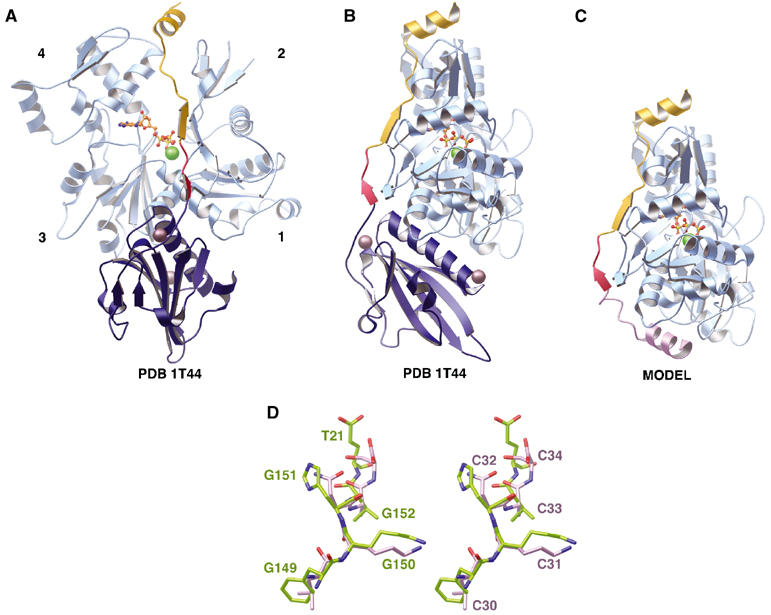
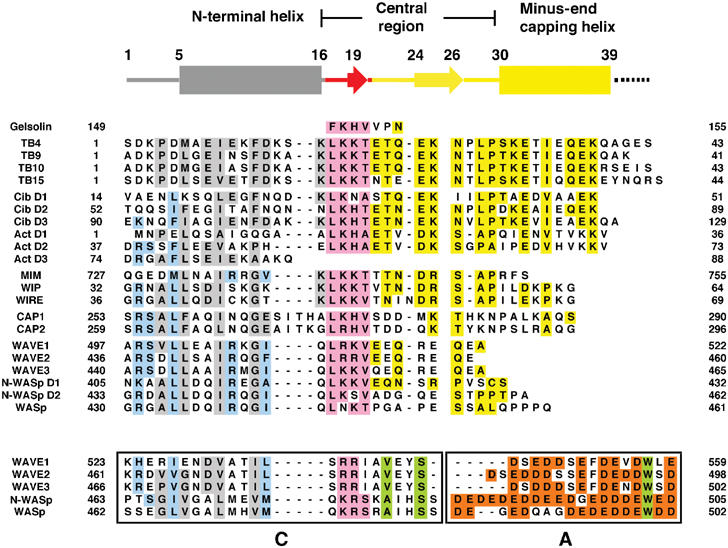
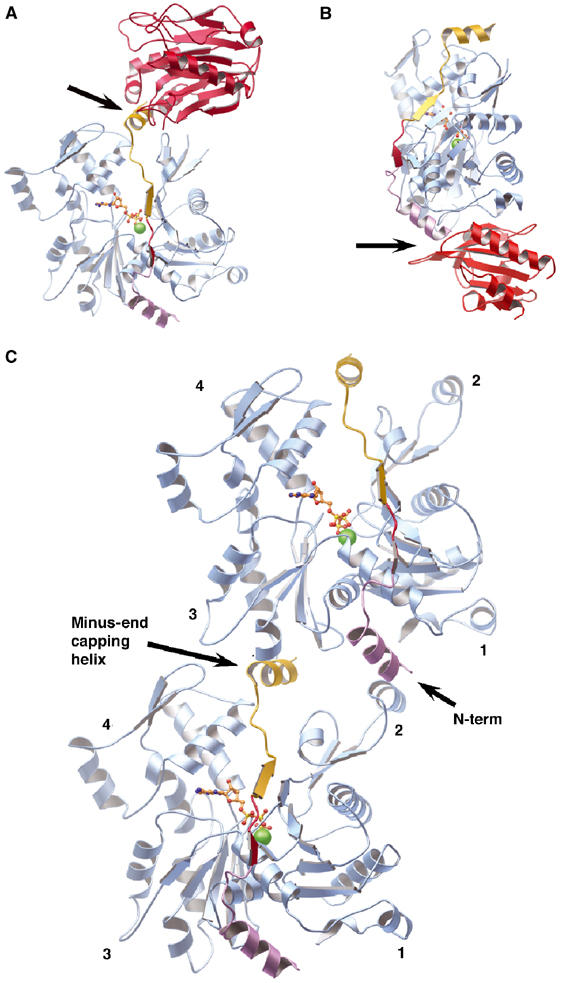
The WH2 (Wiscott-Aldridge syndrome protein homology domain 2) repeat is an actin interacting motif found in monomer sequestering and filament assembly proteins. We have stabilized the prototypical WH2 family member, thymosin-beta4 (Tbeta4), with respect to actin, by creating a hybrid between gelsolin domain 1 and the C-terminal half of Tbeta4 (G1-Tbeta4). This hybrid protein sequesters actin monomers, severs actin filaments and acts as a leaky barbed end cap. Here, we present the structure of the G1-Tbeta4:actin complex at 2 A resolution. The structure reveals that Tbeta4 sequesters by capping both ends of the actin monomer, and that exchange of actin between Tbeta4 and profilin is mediated by a minor overlap in binding sites. The structure implies that multiple WH2 motif-containing proteins will associate longitudinally with actin filaments. Finally, we discuss the role of the WH2 motif in arp2/3 activation.
Figures

Similar articles
-
The structural basis of actin interaction with multiple WH2/beta-thymosin motif-containing proteins.Structure. 2006 Mar;14(3):469-76. doi: 10.1016/j.str.2005.12.011. Structure. 2006. PMID: 16531231
-
The beta-thymosin/WH2 fold: multifunctionality and structure.Ann N Y Acad Sci. 2007 Sep;1112:86-94. doi: 10.1196/annals.1415.011. Epub 2007 Apr 27. Ann N Y Acad Sci. 2007. PMID: 17468236 Review.
-
Structure, function, and evolution of the beta-thymosin/WH2 (WASP-Homology2) actin-binding module.Ann N Y Acad Sci. 2007 Sep;1112:67-75. doi: 10.1196/annals.1415.037. Ann N Y Acad Sci. 2007. PMID: 17947587 Review.
-
Actin-bound structures of Wiskott-Aldrich syndrome protein (WASP)-homology domain 2 and the implications for filament assembly.Proc Natl Acad Sci U S A. 2005 Nov 15;102(46):16644-9. doi: 10.1073/pnas.0507021102. Epub 2005 Nov 7. Proc Natl Acad Sci U S A. 2005. PMID: 16275905 Free PMC article.
-
How a single residue in individual β-thymosin/WH2 domains controls their functions in actin assembly.EMBO J. 2012 Feb 15;31(4):1000-13. doi: 10.1038/emboj.2011.461. Epub 2011 Dec 23. EMBO J. 2012. PMID: 22193718 Free PMC article.
Cited by
-
A Tobacco-Derived Thymosin β4 Concatemer Promotes Cell Proliferation and Wound Healing in Mice.Biomed Res Int. 2016;2016:1973413. doi: 10.1155/2016/1973413. Epub 2016 Jul 14. Biomed Res Int. 2016. PMID: 27493953 Free PMC article.
-
Structure of a longitudinal actin dimer assembled by tandem w domains: implications for actin filament nucleation.J Mol Biol. 2010 Oct 15;403(1):11-23. doi: 10.1016/j.jmb.2010.08.040. Epub 2010 Sep 8. J Mol Biol. 2010. PMID: 20804767 Free PMC article.
-
Identification of Wiskott-Aldrich syndrome protein (WASP) binding sites on the branched actin filament nucleator Arp2/3 complex.Proc Natl Acad Sci U S A. 2018 Feb 13;115(7):E1409-E1418. doi: 10.1073/pnas.1716622115. Epub 2018 Jan 31. Proc Natl Acad Sci U S A. 2018. PMID: 29386393 Free PMC article.
-
Emerging Functions of Actins and Actin Binding Proteins in Trypanosomatids.Front Cell Dev Biol. 2020 Oct 9;8:587685. doi: 10.3389/fcell.2020.587685. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33163497 Free PMC article. Review.
-
Structural characterization of a capping protein interaction motif defines a family of actin filament regulators.Nat Struct Mol Biol. 2010 Apr;17(4):497-503. doi: 10.1038/nsmb.1792. Epub 2010 Mar 28. Nat Struct Mol Biol. 2010. PMID: 20357771 Free PMC article.
References
-
- Ballweber E, Giehl K, Hannappel E, Huff T, Jockusch BM, Mannherz HG (1998) Plant profilin induces actin polymerization from actin:beta-thymosin complexes and competes directly with beta-thymosins and with negative co-operativity with DNase I for binding to actin. FEBS Lett 425: 251–255 - PubMed
-
- Ballweber E, Hannappel E, Huff T, Stephan H, Haener M, Taschner N, Stoffler D, Aebi U, Mannherz HG (2002) Polymerisation of chemically cross-linked actin:thymosin beta(4) complex to filamentous actin: alteration in helical parameters and visualisation of thymosin beta(4) binding on F-actin. J Mol Biol 315: 613–625 - PubMed
-
- Blanchoin L, Amann KJ, Higgs HN, Marchand JB, Kaiser DA, Pollard TD (2000) Direct observation of dendritic actin filament networks nucleated by Arp2/3 complex and WASP/Scar proteins. Nature 404: 1007–1011 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
