

Human cytomegalovirus UL76 encodes a novel virion-associated protein that is able to inhibit viral replication
- PMID: 15331708
- PMCID: PMC515012
- DOI: 10.1128/JVI.78.18.9750-9762.2004
Human cytomegalovirus UL76 encodes a novel virion-associated protein that is able to inhibit viral replication
Abstract
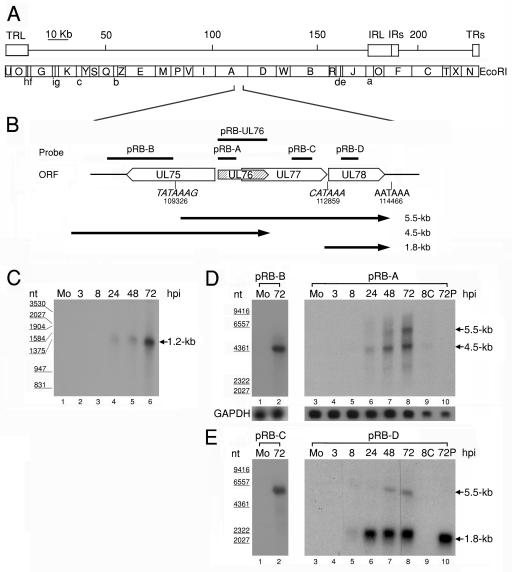
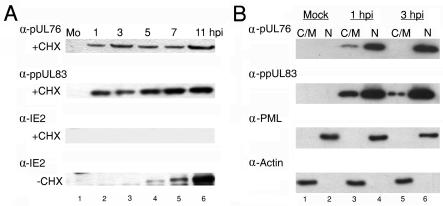
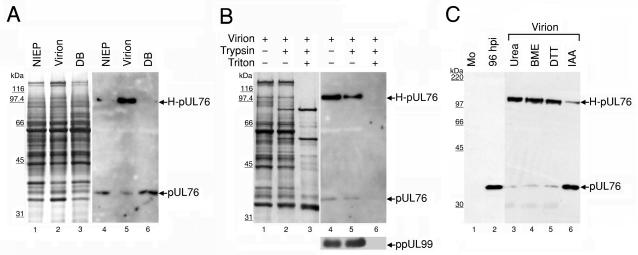
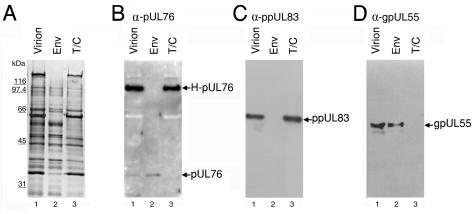
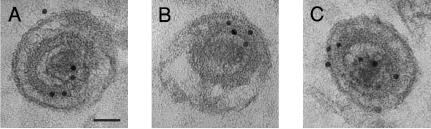
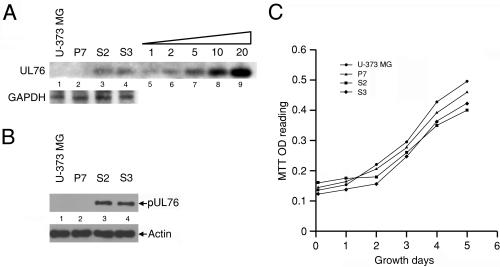
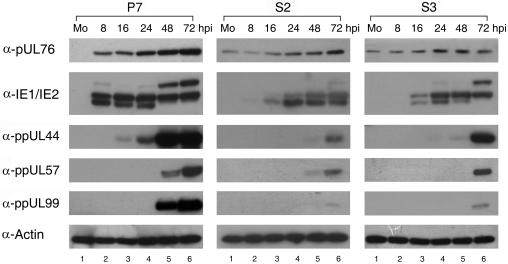
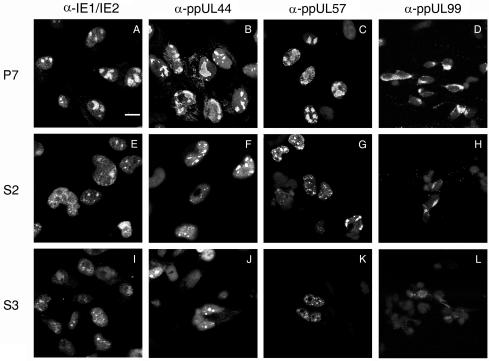
The human cytomegalovirus (HCMV) UL76 gene encodes a highly conserved herpesvirus protein, pUL76, which is able to modulate gene expression in either activation or repression. In this study, two specific transcripts were found to contain the reading frame of UL76, one a 4.5-kb and the other a 5.5-kb tricistronic mRNA encoding the UL76, UL77, and UL78 open reading frames. Both transcripts were expressed with true late kinetics, as revealed by data showing inhibition of production in the presence of phosphonoformic acid. Immediately after viral infection, pUL76 was found in the nuclear fraction and was detected in cells in the presence of the protein synthesis inhibitor cycloheximide. Subsequent virus particle purification and Western blot analysis revealed that two forms of pUL76 are associated within mature virions. The high-molecular-mass protein (H-pUL76) was verified as originating from a free form of pUL76 by cross-linking with an unknown protein(s). By performing a biochemical fractionation experiment with purified virions, we provide evidence that pUL76 and H-pUL76 are associated with the detergent-soluble (envelope) and -insoluble (tegument/capsid) fractions, respectively. Both results were consistent with the images exhibited by immunoelectron microscopy, which showed that the distribution of gold particles labeled by the anti-pUL76 antibody juxtaposed the compartments of the envelope and the tegument/capsid of the virion. Evidence indicated that expression of pUL76 at the immediate-early phase of the viral replication cycle leads to the inhibition of HCMV production. The viral constituent pUL76, with a dominant-negative effect on replication, may provide a novel mechanism for HCMV's resumption of latency.
Copyright 2004 American Society for Microbiology
Figures

References
-
- Bhella, D., F. J. Rixon, and D. J. Dargan. 2000. Cryomicroscopy of human cytomegalovirus virions reveals more densely packed genomic DNA than in herpes simplex virus type 1. J. Mol. Biol. 295:155-161. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
