

Interactions of foot-and-mouth disease virus with soluble bovine alphaVbeta3 and alphaVbeta6 integrins
- PMID: 15331710
- PMCID: PMC514961
- DOI: 10.1128/JVI.78.18.9773-9781.2004
Interactions of foot-and-mouth disease virus with soluble bovine alphaVbeta3 and alphaVbeta6 integrins
Abstract
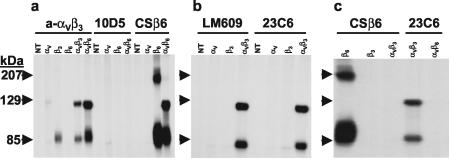
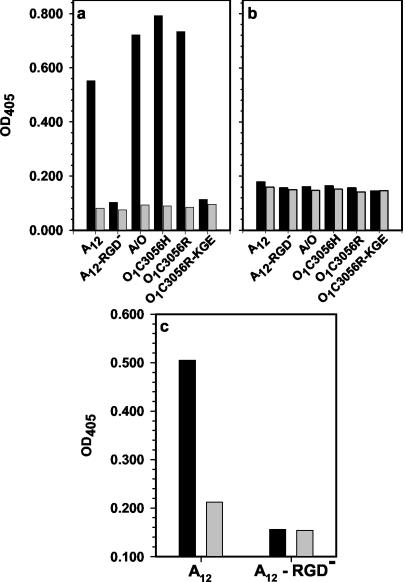
At least four members of the integrin family of receptors, alphaVbeta1, alphaVbeta3, alphaVbeta6, and alphaVbeta8, have been identified as receptors for foot-and-mouth disease virus (FMDV) in vitro. Our investigators have recently shown that the efficiency of receptor usage appears to be related to the viral serotype and may be influenced by structural differences on the viral surface (H. Duque and B. Baxt, J. Virol. 77:2500-2511, 2003). To further examine these differences, we generated soluble alphaVbeta3 and alphaVbeta6 integrins. cDNA plasmids encoding the individual complete integrin alphaV, beta3, and beta6 subunits were used to amplify sequences encoding the subunits' signal peptide and ectodomain, resulting in subunits lacking transmembrane and cytoplasmic domains. COS-1 cells were transfected with plasmids encoding the soluble alphaV subunit and either the soluble beta3 or beta6 subunit and labeled with [35S]methionine-cysteine. Complete subunit heterodimeric integrins were secreted into the medium, as determined by radioimmunoprecipitation with specific monoclonal and polyclonal antibodies. For the examination of the integrins' biological activities, stable cell lines producing the soluble integrins were generated in HEK 293A cells. In the presence of divalent cations, soluble alphaVbeta6 bound to representatives of type A or O viruses, immobilized on plastic dishes, and significantly inhibited viral replication, as determined by plaque reduction assays. In contrast, soluble alphaVbeta3 was unable to bind to immobilized virus of either serotype; however, virus bound to the immobilized integrin, suggesting that FMDV binding to alphaVbeta3 is a low-affinity interaction. In addition, soluble alphaVbeta3 did not neutralize virus infectivity. Incubation of soluble alphaVbeta6 with labeled type A12 or O1 resulted in a significant inhibition of virus adsorption to BHK cells, while soluble alphaVbeta3 caused a low (20 to 30%), but consistent, inhibition of virus adsorption. Virus incubated with soluble alphaVbeta6 had a lower sedimentation rate than native virus on sucrose density gradients, but the particles retained all of their structural proteins and still contained bound integrin and, therefore, were not exhibiting characteristics of a picornavirus A particle.
Copyright 2004 American Society for Microbiology
Figures

Similar articles
-
Molecular characterization and expression analysis of porcine integrins alphavbeta3, alphavbeta6 and alphavbeta8 that are potentially involved in FMDV infection.Mol Cell Probes. 2010 Oct;24(5):256-65. doi: 10.1016/j.mcp.2010.04.005. Epub 2010 May 8. Mol Cell Probes. 2010. PMID: 20438833
-
Sheep (Ovis aries) integrins alphavbeta1 and alphavbeta6 related to foot-and-mouth disease virus infection: molecular cloning, sequence analysis and comparison with homologues.Mol Cell Probes. 2009 Oct;23(5):247-57. doi: 10.1016/j.mcp.2009.06.001. Epub 2009 Jun 23. Mol Cell Probes. 2009. PMID: 19555755
-
Integrin alphaVbeta6 is a high-affinity receptor for coxsackievirus A9.J Gen Virol. 2009 Jan;90(Pt 1):197-204. doi: 10.1099/vir.0.004838-0. J Gen Virol. 2009. PMID: 19088289
-
Cellular receptors for foot and mouth disease virus.Intervirology. 2009;52(4):201-12. doi: 10.1159/000226121. Epub 2009 Jun 24. Intervirology. 2009. PMID: 19556802 Review.
-
The roles of integrin αvβ6 in cancer.Cancer Lett. 2017 Sep 10;403:128-137. doi: 10.1016/j.canlet.2017.06.012. Epub 2017 Jun 17. Cancer Lett. 2017. PMID: 28634043 Review.
Cited by
-
The role of integrins in cancer and the development of anti-integrin therapeutic agents for cancer therapy.Perspect Medicin Chem. 2008 Apr 10;2:57-73. Perspect Medicin Chem. 2008. PMID: 19787098 Free PMC article.
-
Picornaviruses.Curr Top Microbiol Immunol. 2010;343:43-89. doi: 10.1007/82_2010_37. Curr Top Microbiol Immunol. 2010. PMID: 20397067 Free PMC article.
-
Heparan sulfate-binding foot-and-mouth disease virus enters cells via caveola-mediated endocytosis.J Virol. 2008 Sep;82(18):9075-85. doi: 10.1128/JVI.00732-08. Epub 2008 Jul 9. J Virol. 2008. PMID: 18614639 Free PMC article.
-
Early events in integrin alphavbeta6-mediated cell entry of foot-and-mouth disease virus.J Virol. 2005 Jul;79(13):8519-34. doi: 10.1128/JVI.79.13.8519-8534.2005. J Virol. 2005. PMID: 15956594 Free PMC article.
-
How foot-and-mouth disease virus receptor mediates foot-and-mouth disease virus infection.Virol J. 2015 Feb 3;12:9. doi: 10.1186/s12985-015-0246-z. Virol J. 2015. PMID: 25645358 Free PMC article. Review.
References
-
- Acharya, R., E. Fry, D. Stuart, G. Fox, D. Rowlands, and F. Brown. 1989. The three-dimensional structure of foot-and-mouth disease virus at 2.9 Å resolution. Nature 337:709-716. - PubMed
-
- Bachrach, H. L. 1968. Foot-and-mouth disease. Annu. Rev. Microbiol. 22:201-244. - PubMed
-
- Bachrach, H. L., R. Trautman, and S. S. Breese. 1964. Chemical and physical properties of virtually pure foot-and-mouth disease virus. Am. J. Vet. Res. 25:333-342. - PubMed
-
- Baranowski, E., C. M. Ruiz-Jarabo, and E. Domingo. 2001. Evolution of cell recognition by viruses. Science 292:1102-1105. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
