

doi: 10.1083/jcb.200312072.
Membrane insertion of anthrax protective antigen and cytoplasmic delivery of lethal factor occur at different stages of the endocytic pathway
Affiliations
- PMID: 15337774
- PMCID: PMC2172425
- DOI: 10.1083/jcb.200312072
Item in Clipboard
Membrane insertion of anthrax protective antigen and cytoplasmic delivery of lethal factor occur at different stages of the endocytic pathway
J Cell Biol.
.
Abstract
The protective antigen (PA) of anthrax toxin binds to a cell surface receptor, undergoes heptamerization, and binds the enzymatic subunits, the lethal factor (LF) and the edema factor (EF). The resulting complex is then endocytosed. Via mechanisms that depend on the vacuolar ATPase and require membrane insertion of PA, LF and EF are ultimately delivered to the cytoplasm where their targets reside. Here, we show that membrane insertion of PA already occurs in early endosomes, possibly only in the multivesicular regions, but that subsequent delivery of LF to the cytoplasm occurs preferentially later in the endocytic pathway and relies on the dynamics of internal vesicles of multivesicular late endosomes.
Figures
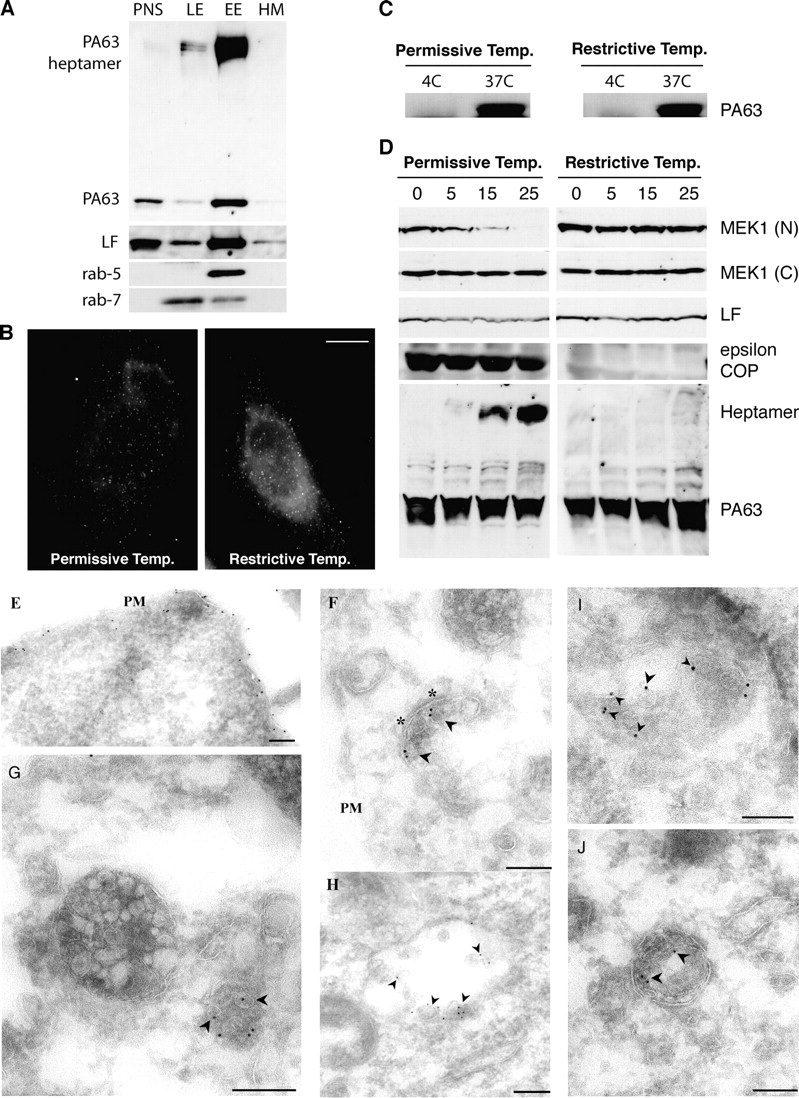
Conversion of PAheptamer to an SDS-resistant form occurs in early endosomes and is COPI dependent. (A) BHK cells were incubated at 4°C, 1 h with 500 ng/ml trypsin-nicked PA (PAn; Abrami et al., 2003) and 20 ng/ml LF, transferred to a toxin-free medium (37°C,1 h) and treated with 5 U/ml trypsin,10 min 37°C, to remove surface-bound toxin. Subcellular fractionation of postnuclear supernatants (PNS) was performed to separate early (EE) from late endosomes (LE) and heavy membranes (HM). 15 μg of protein from each fraction were analyzed by Western blotting for the presence of PA and PAheptamer, LF, rab5, and rab7. (B–D) LdlF cells were grown at the permissive (34°C) or restrictive temperature (40°C) for 18 h. (B) Cells were incubated for 20 min at 4°C with 500 ng/ml PASNKE, a PA variant with a mutated furin cleavage site, submitted to antibody cross-linking at 4°C and further incubated at 37°C for 30 min. Surface-bound toxin was removed by an acid wash (which lead to the observed background staining). Bar, 10 μm. (C) Cells were incubated with 500 ng/ml PAn, 1 h 4°C, washed and incubated at 4°C or 37°C with a toxin-free medium for 15 min. Surface toxin was removed as in A. 40 μg of PNS was analyzed by a SDS-PAGE gel followed by Western blotting against PA. (D) ldlF CHO cells were incubated at 4°C for 1 h with 500 ng/ml PAn and 500 ng/ml LF, transferred to 37°C for different periods of time (in min) in a toxin-free medium. 40 μg of total cell extracts were analyzed by Western blotting to detect LF processed MEK1 (anti–NH2-terminal antibody), total MEK1 (anti–COOH-terminal antibody), LF, ε-COP, and PA. (E–J) BHK cells were incubated with 10 μg/ml PAn for 1 h at 4°C, warmed to 37°C for either 10 min (E) or 40 min (F–J) before fixation and preparation of ultrathin sections. Sections were labeled with rabbit anti-PA followed by 10 nm protein A gold. Labeling is evident on the plasma membrane (E, PM) and in putative early endosomal structures (F), recognized by the presence of a characteristic patch of coat material (asterisks). At later times (G–J) labeling is evident within a subset of multivesicular endosomal structures with enrichment on internal vesicles (arrowheads). Bars, 200 nm.
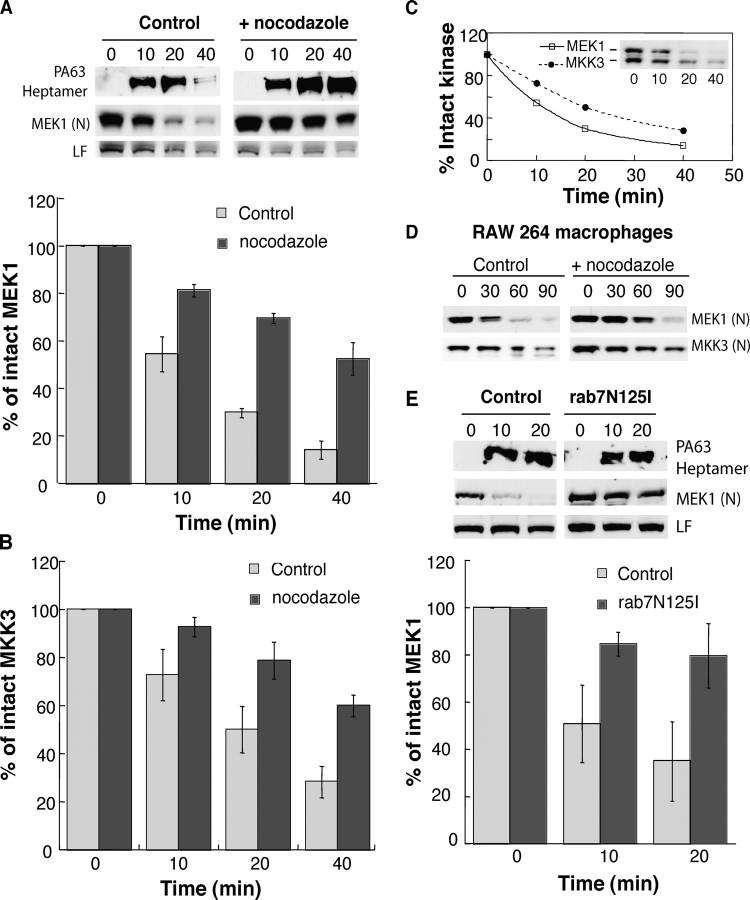
Efficient encounter between LF and its target occurs from late endosomes. (A) CHO cells were incubated or not with 10 μM nocodazole for 2 h at 37°C (nocodazole was then present throughout the entire experiment), followed by 1 h at 4°C with 500 ng/ml PAn and 20 ng/ml LF, transferred to 37°C for different periods of time (in min) in a toxin-free medium. 40 μg of PNS was analyzed by Western blotting against PA, the NH2 terminus of MEK1 (N) and LF. The amount of intact MEK1 (A) and MKK3 (B) was quantified by densitometry and normalized to the amount at time t = 0 (n = 4, errors bars represent SDs). (C) The kinetics of decrease in intact MEK1 were compared with that of decrease in MKK3. Western blots (inset) were quantified by densitometry as in A and B. (D) RAW 264 macrophages were incubated or not with 10 μM nocodazole (2 h at 37°C), followed by 1 h at 4°C with 500 ng/ml PAn and 100 ng/ml LF. 40 μg of PNS was analyzed by Western blotting against the NH2 termini of MEK1 and MKK3. (E) HeLa cells were transfected or not with dominant-negative rab7N125I cDNA, incubated at 4°C for 1 h with 500 ng/ml PAn and 500 ng/ml LF and transferred to 37°C for different periods of time (in min) in a toxin-free medium. 40 μg of PNS was analyzed as in A (n = 4).
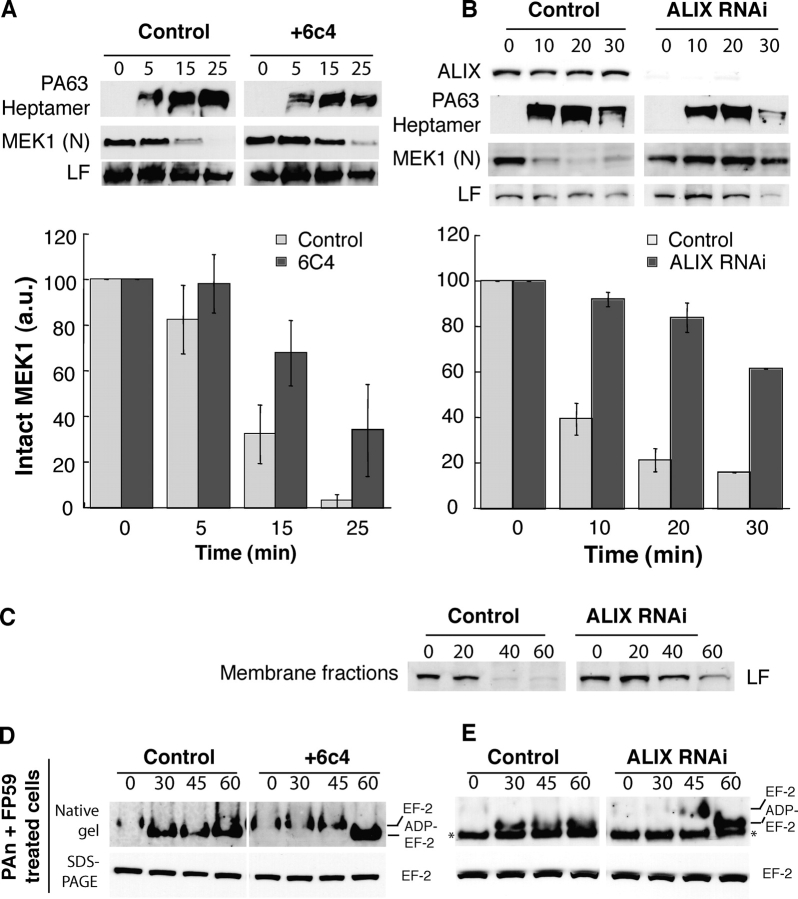
Alteration of the dynamics of late endosomal intraluminal vesicles leads to a delay in MEK1 cleavage by LF. (A) CHO cells were incubated or not for 18 h with the anti-LBPA antibody 6c4 (50 μg/ml), washed, further incubated at 4°C for 1 h with 500 ng/ml PAn and 250 ng/ml LF, and transferred to 37°C for different periods of time (in min) in a toxin-free medium. 20 μg of PNS was analyzed by Western blotting to detect PAheptamer, the NH2 terminus of MEK1 and LF. The amount of intact MEK1 was quantified as in Fig. 2 A (n = 4). (B) HeLa cells were transfected with siRNA against ALIX, the efficiency of which was examined 78 h later by Western blotting using antibodies against ALIX. Cells were then incubated at 4°C for 1 h with 500 ng/ml PAn and 100 ng/ml LF, transferred to 37°C for different periods of time (in min) in a toxin-free medium, homogenized, and 20 μg of PNS was analyzed to detect the presence of PAheptamer and the NH2 terminus of MEK1. Quantifications were performed as in A. (C) HeLa cells were treated as in B. 80 μg of PNS were centrifuged at 100,000 g for 1 h and the pellet analyzed by Western blotting for LF. (D) CHO cells were treated as in A and incubated with 500 ng/ml PAn and 500 ng/ml FP59. 20 μg of cell lysates were loaded on native or SDS-PAGE before Western blotting against EF-2. (E) ALIX was knocked down in HeLa cells as in B, incubated with 500 ng/ml PAn with 500 ng/ml FP59, and the modification of EF2 analyzed as in D.
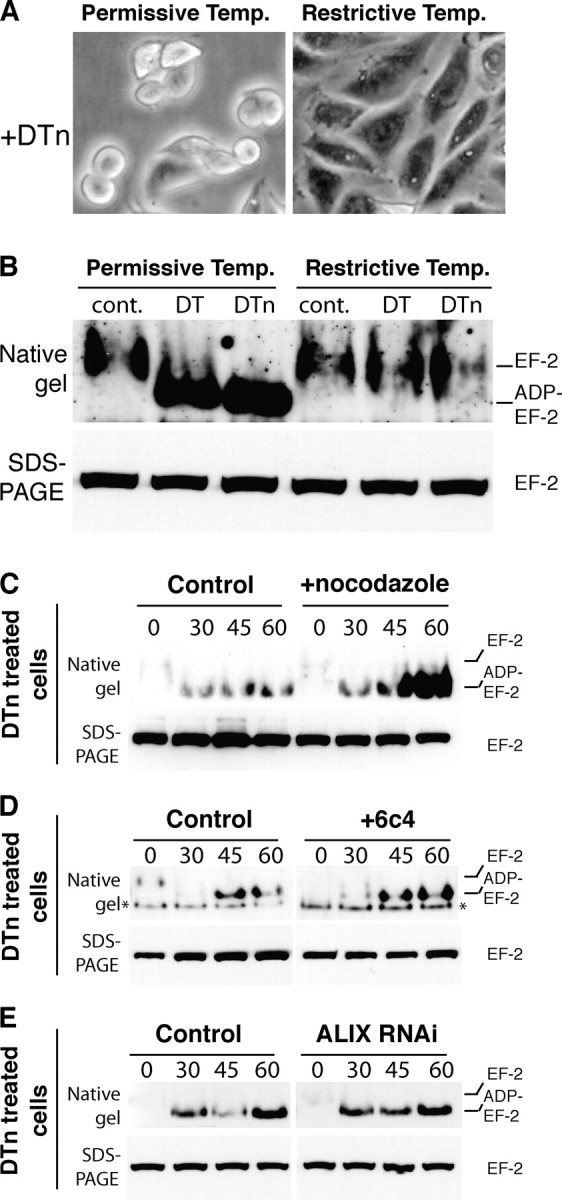
Cytosolic delivery of DT is COPI dependent but does not require transport to late endosomes. (A and B) ldlF cells were grown at the permissive or restrictive temperature for 16 h, incubated for 7 h (A) or 2.5 h (B) at 37°C in serum-free medium with 500 ng/ml of DT or trypsin-nicked DT (DTn) or left untreated (cont.). (B) Cell lysates were prepared and the modification of EF2 analyzed as in Fig. 3 D. (C) Wild-type CHO cells were incubated or not with 10 μM nocodazole for 2 h at 37°C and 15 min at 4°C, followed by 1 h at 4°C with 500 ng/ml DTn, washed and further incubated for different times at 37°C. The modification of EF2 was analyzed as above. (D) CHO cells were treated with 6c4 antibody as in Fig. 3 A and incubated with 500 ng/ml DTn and modification of EF2 analyzed. The band labeled with an asterisk is a cross-reacting protein, apparent in certain experiments. (E) ALIX was knocked down in HeLa cells as in Fig. 3 B, incubated with 500 ng/ml DTn, and modification of EF2 analyzed.
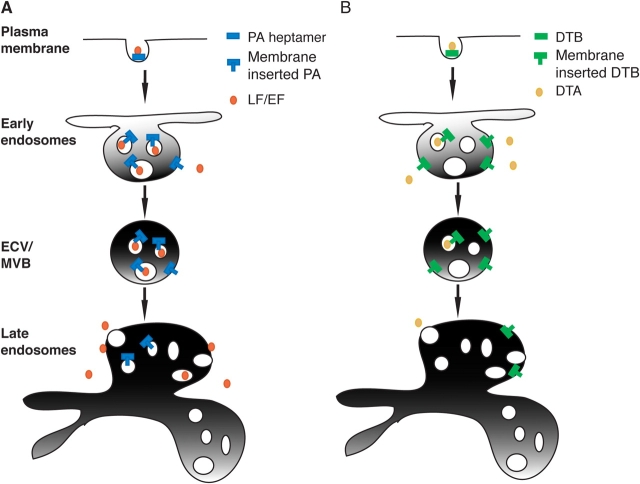
Endocytic routes of anthrax and diphtheria toxins. Based on the present work and the literature, we propose the following models for the delivery of anthrax lethal toxin (A) and DT (B) to the cytoplasm. Both toxins are internalized, transported to early endosomes, and sorted into the vesicular regions. (A) Anthrax toxin would be sorted into intraluminal vesicles whereby membrane insertion of PA and translocation of LF would result in trapping of the metalloprotease in the lumen of intraluminal vesicles. Once transported to late endosomes, back fusion of intraluminal vesicles with the limiting membrane would deliver LF to the cytoplasm. (B) In contrast, in early endosomes, DT would preferentially insert into the limiting membrane allowing direct delivery to the cytoplasm.
References
-
- Abrami, L., M. Fivaz, E. Decroly, N.G. Seidah, J. François, G. Thomas, S. Leppla, J.T. Buckley, and F.G. van der Goot. 1998. The pore-forming toxin proaerolysin is processed by furin. J. Biol. Chem. 273:32656–32661. - PubMed
-
- Chow, A., D. Toomre, W. Garrett, and I. Mellman. 2002. Dendritic cell maturation triggers retrograde MHC class II transport from lysosomes to the plasma membrane. Nature. 418:988–994. - PubMed
-
- Collier, R.J., and J.A. Young. 2003. Anthrax toxin. Annu. Rev. Cell Dev. Biol. 19:45–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
