

Drosophila Ada2b is required for viability and normal histone H3 acetylation
- PMID: 15340070
- PMCID: PMC515027
- DOI: 10.1128/MCB.24.18.8080-8089.2004
Drosophila Ada2b is required for viability and normal histone H3 acetylation
Abstract
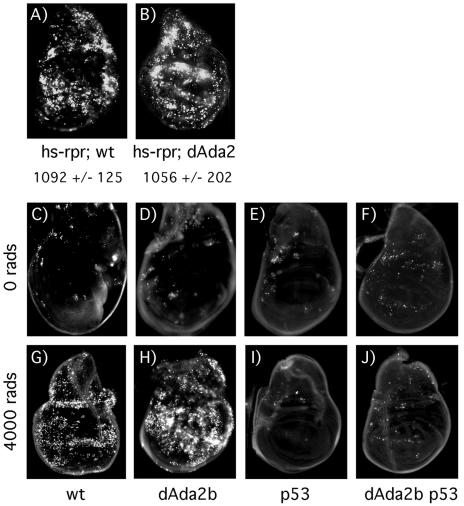
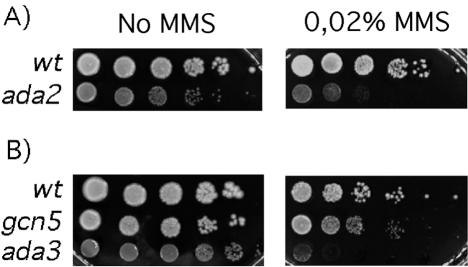
Regulation of chromatin through histone acetylation is an important step in gene expression. The Gcn5 histone acetyltransferase is part of protein complexes, e.g., the SAGA complex, that interact with transcriptional activators, targeting the enzyme to specific promoters and assisting in recruitment of the basal RNA polymerase transcription machinery. The Ada2 protein directly binds to Gcn5 and stimulates its catalytic activity. Drosophila contains two Ada2 proteins, Drosophila Ada2a (dAda2a) and dAda2b. We have generated flies that lack dAda2b, which is part of a Drosophila SAGA-like complex. dAda2b is required for viability in Drosophila, and its deletion causes a reduction in histone H3 acetylation. A global hypoacetylation of chromatin was detected on polytene chromosomes in dAda2b mutants. This indicates that the dGcn5-dAda2b complex could have functions in addition to assisting in transcriptional activation through gene-specific acetylation. Although the Drosophila p53 protein was previously shown to interact with the SAGA-like complex in vitro, we find that p53 induction of reaper gene expression occurs normally in dAda2b mutants. Moreover, dAda2b mutant animals show excessive p53-dependent apoptosis in response to gamma radiation. Based on this result, we speculate that dAda2b may be necessary for efficient DNA repair or generation of a DNA damage signal. This could be an evolutionarily conserved function, since a yeast ada2 mutant is also sensitive to a genotoxic agent.
Figures

Similar articles
-
Two different Drosophila ADA2 homologues are present in distinct GCN5 histone acetyltransferase-containing complexes.Mol Cell Biol. 2003 Jan;23(1):306-21. doi: 10.1128/MCB.23.1.306-321.2003. Mol Cell Biol. 2003. PMID: 12482983 Free PMC article.
-
The Drosophila histone acetyltransferase Gcn5 and transcriptional adaptor Ada2a are involved in nucleosomal histone H4 acetylation.Mol Cell Biol. 2006 Dec;26(24):9413-23. doi: 10.1128/MCB.01401-06. Epub 2006 Oct 9. Mol Cell Biol. 2006. PMID: 17030603 Free PMC article.
-
Two Drosophila Ada2 homologues function in different multiprotein complexes.Mol Cell Biol. 2003 May;23(9):3305-19. doi: 10.1128/MCB.23.9.3305-3319.2003. Mol Cell Biol. 2003. PMID: 12697829 Free PMC article.
-
The Gcn5 complexes in Drosophila as a model for metazoa.Biochim Biophys Acta Gene Regul Mech. 2021 Feb;1864(2):194610. doi: 10.1016/j.bbagrm.2020.194610. Epub 2020 Jul 28. Biochim Biophys Acta Gene Regul Mech. 2021. PMID: 32735945 Review.
-
Recruitment of chromatin remodelling factors during gene activation via the glucocorticoid receptor N-terminal domain.Biochem Soc Trans. 2000;28(4):410-4. Biochem Soc Trans. 2000. PMID: 10961930 Review.
Cited by
-
Invadolysin acts genetically via the SAGA complex to modulate chromosome structure.Nucleic Acids Res. 2015 Apr 20;43(7):3546-62. doi: 10.1093/nar/gkv211. Epub 2015 Mar 16. Nucleic Acids Res. 2015. PMID: 25779050 Free PMC article.
-
The Transcriptional Coactivator ADA2b Recruits a Structural Maintenance Protein to Double-Strand Breaks during DNA Repair in Plants.Plant Physiol. 2018 Apr;176(4):2613-2622. doi: 10.1104/pp.18.00123. Epub 2018 Feb 20. Plant Physiol. 2018. PMID: 29463775 Free PMC article.
-
Enzymatic modules of the SAGA chromatin-modifying complex play distinct roles in Drosophila gene expression and development.Genes Dev. 2017 Aug 1;31(15):1588-1600. doi: 10.1101/gad.300988.117. Epub 2017 Sep 8. Genes Dev. 2017. PMID: 28887412 Free PMC article.
-
Regulation of presynaptic homeostatic plasticity by glial signalling in Alzheimer's disease.J Physiol. 2024 Dec 20:10.1113/JP286751. doi: 10.1113/JP286751. Online ahead of print. J Physiol. 2024. PMID: 39705214 Review.
-
Transcriptome features of innate immune memory in Drosophila.PLoS Genet. 2022 Oct 17;18(10):e1010005. doi: 10.1371/journal.pgen.1010005. eCollection 2022 Oct. PLoS Genet. 2022. PMID: 36252180 Free PMC article.
References
-
- Aasland, R., A. F. Stewart, and T. Gibson. 1996. The SANT domain: a putative DNA-binding domain in the SWI-SNF and ADA complexes, the transcriptional co-repressor N-CoR and TFIIIB. Trends Biochem. Sci. 21:87-88. - PubMed
-
- Abraham, R. T. 2001. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 15:2177-2196. - PubMed
-
- Barlev, N. A., R. Candau, L. Wang, P. Darpino, N. Silverman, and S. L. Berger. 1995. Characterization of physical interactions of the putative transcriptional adaptor, ADA2, with acidic activation domains and TATA-binding protein. J. Biol. Chem. 270:19337-19344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous