

Transcriptional regulation of human CYP27 integrates retinoid, peroxisome proliferator-activated receptor, and liver X receptor signaling in macrophages
- PMID: 15340076
- PMCID: PMC515045
- DOI: 10.1128/MCB.24.18.8154-8166.2004
Transcriptional regulation of human CYP27 integrates retinoid, peroxisome proliferator-activated receptor, and liver X receptor signaling in macrophages
Abstract
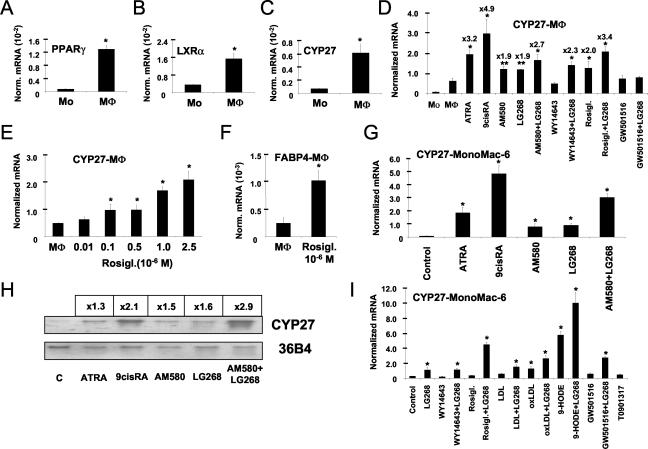
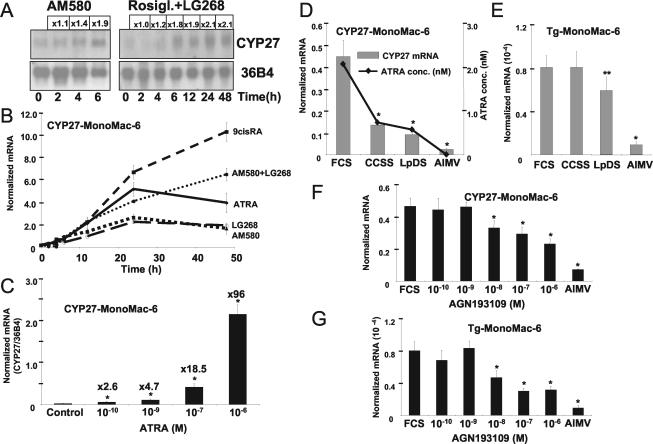
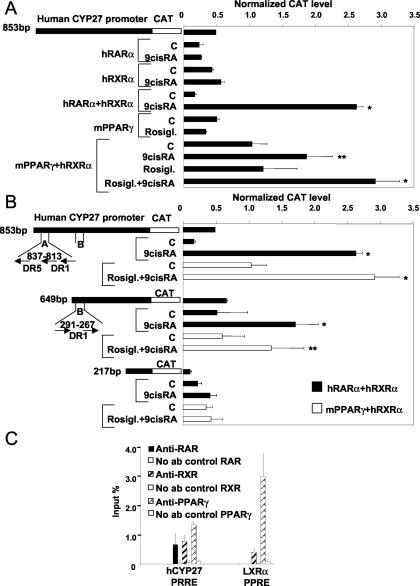
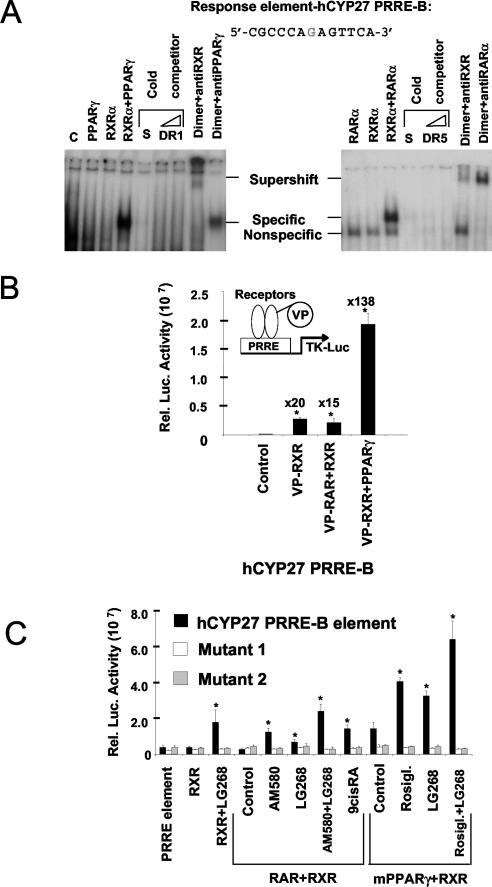
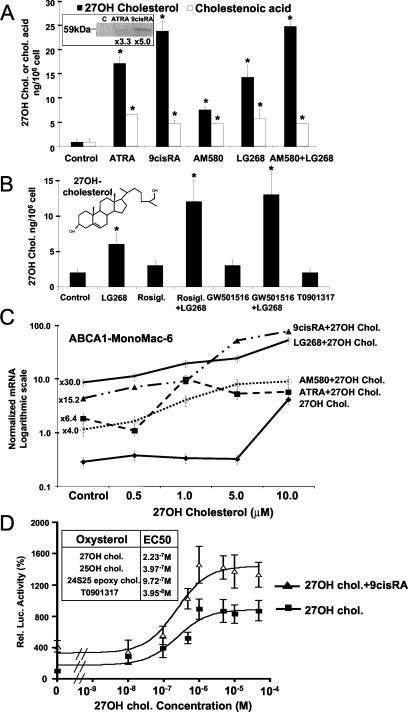
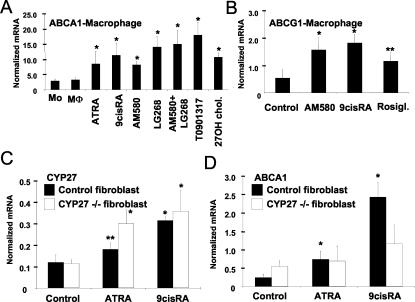
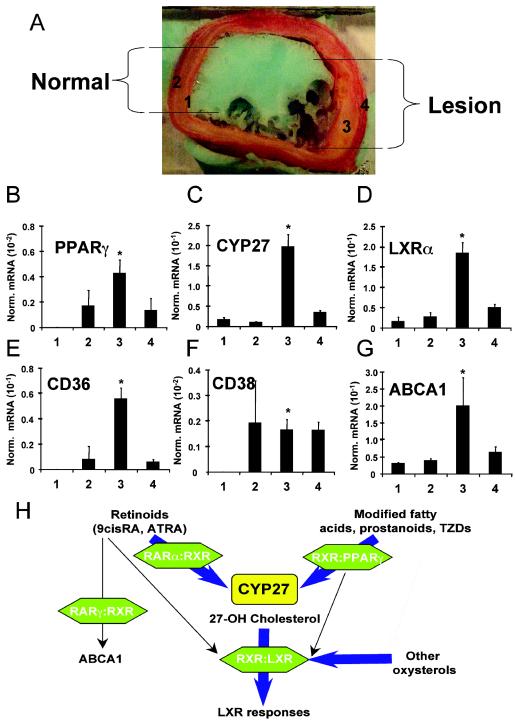
Cholesterol uptake and efflux are key metabolic processes associated with macrophage physiology and atherosclerosis. Peroxisome proliferator-activated receptor gamma (PPARgamma) and liver X receptor alpha (LXRalpha) have been linked to the regulation of these processes. It remains to be identified how activation of these receptors is connected and regulated by endogenous lipid molecules. We identified CYP27, a p450 enzyme, as a link between retinoid, PPARgamma, and LXR signaling. We show that the human CYP27 gene is under coupled regulation by retinoids and ligands of PPARs via a PPAR-retinoic acid receptor response element in its promoter. Induction of the enzyme's expression results in an increased level of 27-hydroxycholesterol and upregulation of LXR-mediated processes. Upregulated CYP27 activity also leads to LXR-independent elimination of CYP27 metabolites as an alternative means of cholesterol efflux. Moreover, human macrophage-rich atherosclerotic lesions have an increased level of retinoid-, PPARgamma-, and LXR-regulated gene expression and also enhanced CYP27 levels. Our findings suggest that nuclear receptor-regulated CYP27 expression is likely to be a key integrator of retinoic acid receptor-PPARgamma-LXR signaling, relying on natural ligands and contributing to lipid metabolism in macrophages.
Figures
Similar articles
-
Cross-talk between peroxisome proliferator-activated receptor (PPAR) alpha and liver X receptor (LXR) in nutritional regulation of fatty acid metabolism. II. LXRs suppress lipid degradation gene promoters through inhibition of PPAR signaling.Mol Endocrinol. 2003 Jul;17(7):1255-67. doi: 10.1210/me.2002-0191. Epub 2003 May 1. Mol Endocrinol. 2003. PMID: 12730332
-
27-hydroxycholesterol is an endogenous ligand for liver X receptor in cholesterol-loaded cells.J Biol Chem. 2001 Oct 19;276(42):38378-87. doi: 10.1074/jbc.M105805200. Epub 2001 Aug 14. J Biol Chem. 2001. PMID: 11504730
-
Cross-talk between peroxisome proliferator-activated receptor (PPAR) alpha and liver X receptor (LXR) in nutritional regulation of fatty acid metabolism. I. PPARs suppress sterol regulatory element binding protein-1c promoter through inhibition of LXR signaling.Mol Endocrinol. 2003 Jul;17(7):1240-54. doi: 10.1210/me.2002-0190. Epub 2003 May 1. Mol Endocrinol. 2003. PMID: 12730331
-
The role of lipid-activated nuclear receptors in shaping macrophage and dendritic cell function: From physiology to pathology.J Allergy Clin Immunol. 2013 Aug;132(2):264-86. doi: 10.1016/j.jaci.2013.05.044. J Allergy Clin Immunol. 2013. PMID: 23905916 Review.
-
Oxysterol signaling links cholesterol metabolism and inflammation via the liver X receptor in macrophages.Mol Aspects Med. 2009 Jun;30(3):134-52. doi: 10.1016/j.mam.2009.02.002. Epub 2009 Feb 25. Mol Aspects Med. 2009. PMID: 19248804 Review.
Cited by
-
Arginine methylation provides epigenetic transcription memory for retinoid-induced differentiation in myeloid cells.Mol Cell Biol. 2005 Jul;25(13):5648-63. doi: 10.1128/MCB.25.13.5648-5663.2005. Mol Cell Biol. 2005. PMID: 15964820 Free PMC article.
-
Local and systemic gene expression responses of Atlantic salmon (Salmo salar L.) to infection with the salmon louse (Lepeophtheirus salmonis).BMC Genomics. 2008 Oct 23;9:498. doi: 10.1186/1471-2164-9-498. BMC Genomics. 2008. PMID: 18945374 Free PMC article.
-
Liver x receptor: a novel therapeutic target.Indian J Pharm Sci. 2008 Mar-Apr;70(2):135-44. doi: 10.4103/0250-474X.41445. Indian J Pharm Sci. 2008. PMID: 20046702 Free PMC article.
-
The Molecular and Genetic Interactions between Obesity and Breast Cancer Risk.Medicina (Kaunas). 2023 Jul 21;59(7):1338. doi: 10.3390/medicina59071338. Medicina (Kaunas). 2023. PMID: 37512149 Free PMC article. Review.
-
A Crosstalk Between Brain Cholesterol Oxidation and Glucose Metabolism in Alzheimer's Disease.Front Neurosci. 2019 May 31;13:556. doi: 10.3389/fnins.2019.00556. eCollection 2019. Front Neurosci. 2019. PMID: 31213973 Free PMC article. Review.
References
-
- Andersson, S., D. L. Davis, H. Dahlback, H. Jornvall, and D. W. Russell. 1989. Cloning, structure, and expression of the mitochondrial cytochrome P-450 sterol 26-hydroxylase, a bile acid biosynthetic enzyme. J. Biol. Chem. 264:8222-8229. - PubMed
-
- Babiker, A., O. Andersson, D. Lindblom, J. van der Linden, B. Wiklund, D. Lutjohann, U. Diczfalusy, and I. Bjorkhem. 1999. Elimination of cholesterol as cholestenoic acid in human lung by sterol 27-hydroxylase: evidence that most of this steroid in the circulation is of pulmonary origin. J. Lipid Res. 40:1417-1425. - PubMed
-
- Babiker, A., O. Andersson, E. Lund, R. J. Xiu, S. Deeb, A. Reshef, E. Leitersdorf, U. Diczfalusy, and I. Bjorkhem. 1997. Elimination of cholesterol in macrophages and endothelial cells by the sterol 27-hydroxylase mechanism. Comparison with high density lipoprotein-mediated reverse cholesterol transport. J. Biol. Chem. 272:26253-26261. - PubMed
-
- Benko, S., J. D. Love, M. Beladi, J. W. Schwabe, and L. Nagy. 2003. Molecular determinants of the balance between co-repressor and co-activator recruitment to the retinoic acid receptor. J. Biol. Chem. 278:43797-43806. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources