

The Swi/Snf chromatin remodeling complex is required for ribosomal DNA and telomeric silencing in Saccharomyces cerevisiae
- PMID: 15340082
- PMCID: PMC515061
- DOI: 10.1128/MCB.24.18.8227-8235.2004
The Swi/Snf chromatin remodeling complex is required for ribosomal DNA and telomeric silencing in Saccharomyces cerevisiae
Abstract
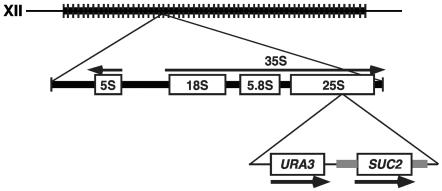
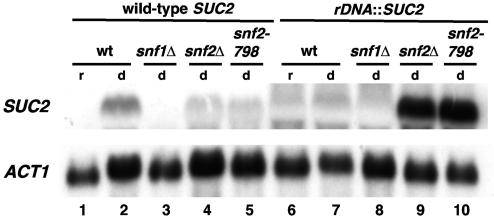
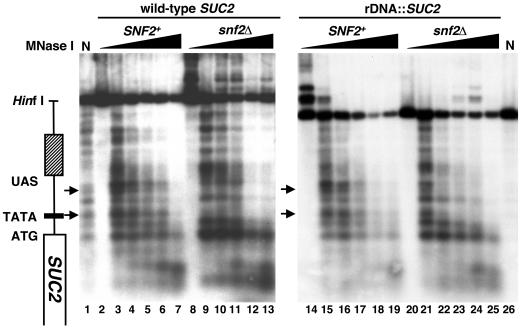
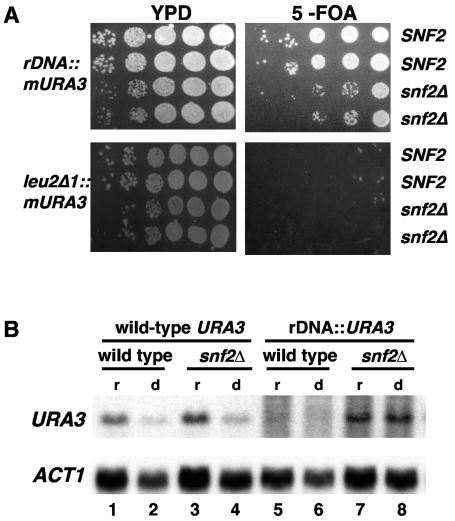
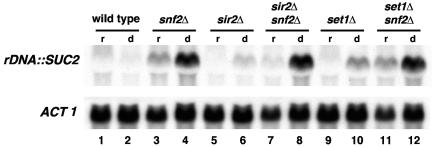
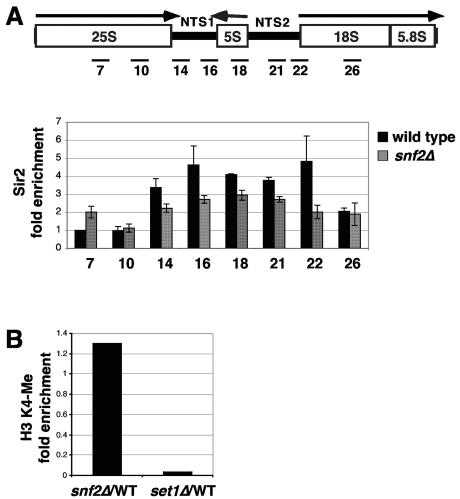
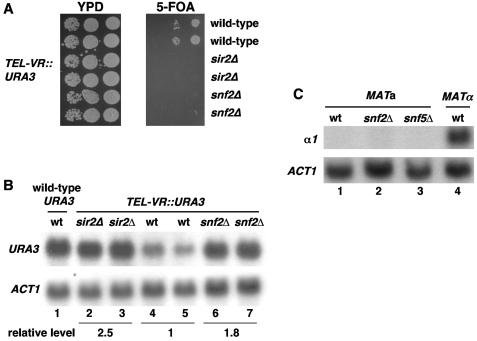
The Swi/Snf chromatin remodeling complex has been previously demonstrated to be required for transcriptional activation and repression of a subset of genes in Saccharomyces cerevisiae. In this work we demonstrate that Swi/Snf is also required for repression of RNA polymerase II-dependent transcription in the ribosomal DNA (rDNA) locus (rDNA silencing). This repression appears to be independent of both Sir2 and Set1, two factors known to be required for rDNA silencing. In contrast to many other rDNA silencing mutants that have elevated levels of rDNA recombination, snf2Delta mutants have a significantly decreased level of rDNA recombination. Additional studies have demonstrated that Swi/Snf is also required for silencing of genes near telomeres while having no detectable effect on silencing of HML or HMR.
Figures
Similar articles
-
Isw1 acts independently of the Isw1a and Isw1b complexes in regulating transcriptional silencing at the ribosomal DNA locus in Saccharomyces cerevisiae.J Mol Biol. 2007 Aug 3;371(1):1-10. doi: 10.1016/j.jmb.2007.04.089. Epub 2007 May 18. J Mol Biol. 2007. PMID: 17561109 Free PMC article.
-
A genetic screen for ribosomal DNA silencing defects identifies multiple DNA replication and chromatin-modulating factors.Mol Cell Biol. 1999 Apr;19(4):3184-97. doi: 10.1128/MCB.19.4.3184. Mol Cell Biol. 1999. PMID: 10082585 Free PMC article.
-
Evidence that Set1, a factor required for methylation of histone H3, regulates rDNA silencing in S. cerevisiae by a Sir2-independent mechanism.Curr Biol. 2002 Jan 22;12(2):165-70. doi: 10.1016/s0960-9822(01)00652-2. Curr Biol. 2002. PMID: 11818070
-
Stay close to your sister.Mol Cell. 2004 May 21;14(4):418-20. doi: 10.1016/s1097-2765(04)00266-7. Mol Cell. 2004. PMID: 15149590 Review.
-
Common themes in mechanisms of gene silencing.Mol Cell. 2001 Sep;8(3):489-98. doi: 10.1016/s1097-2765(01)00340-9. Mol Cell. 2001. PMID: 11583612 Review.
Cited by
-
RNA Sequencing Reveals Specific TranscriptomicSignatures Distinguishing Effects of the [SWI⁺] Prion and SWI1 Deletion in Yeast Saccharomyces cerevisiae.Genes (Basel). 2019 Mar 12;10(3):212. doi: 10.3390/genes10030212. Genes (Basel). 2019. PMID: 30871095 Free PMC article.
-
Fission yeast SWI/SNF and RSC complexes show compositional and functional differences from budding yeast.Nat Struct Mol Biol. 2008 Aug;15(8):873-80. doi: 10.1038/nsmb.1452. Epub 2008 Jul 11. Nat Struct Mol Biol. 2008. PMID: 18622392 Free PMC article.
-
The nuclear pore complex mediates binding of the Mig1 repressor to target promoters.PLoS One. 2011;6(11):e27117. doi: 10.1371/journal.pone.0027117. Epub 2011 Nov 14. PLoS One. 2011. PMID: 22110603 Free PMC article.
-
Stress-dependent dynamics of global chromatin remodeling in yeast: dual role for SWI/SNF in the heat shock stress response.Mol Cell Biol. 2008 Apr;28(7):2221-34. doi: 10.1128/MCB.01659-07. Epub 2008 Jan 22. Mol Cell Biol. 2008. PMID: 18212068 Free PMC article.
-
SWI/SNF and Asf1 independently promote derepression of the DNA damage response genes under conditions of replication stress.PLoS One. 2011;6(6):e21633. doi: 10.1371/journal.pone.0021633. Epub 2011 Jun 27. PLoS One. 2011. PMID: 21738741 Free PMC article.
References
-
- Aparicio, O. M., B. L. Billington, and D. E. Gottschling. 1991. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell 66:1279-1287. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1998. Current protocols in molecular biology. Greene/Wiley-Interscience, New York, N.Y.
-
- Becker, P. B., and W. Horz. 2002. ATP-dependent nucleosome remodeling. Annu. Rev. Biochem. 71:247-273. - PubMed
-
- Brachmann, C. B., A. Davies, G. J. Cost, E. Caputo, J. Li, P. Hieter, and J. D. Boeke. 1998. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14:115-132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases