

Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals
- PMID: 15340154
- PMCID: PMC518791
- DOI: 10.1073/pnas.0402907101
Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals
Abstract
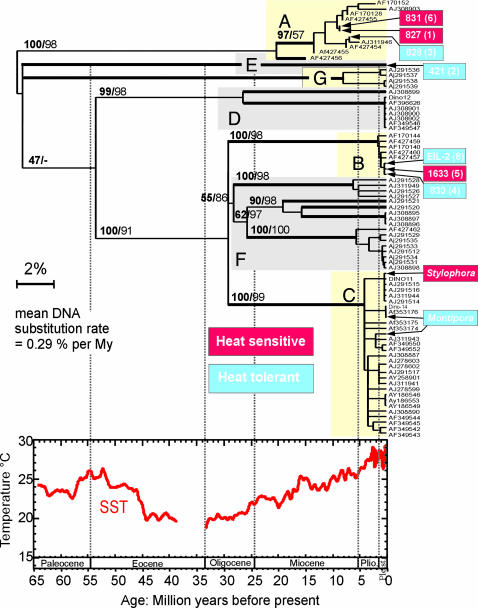
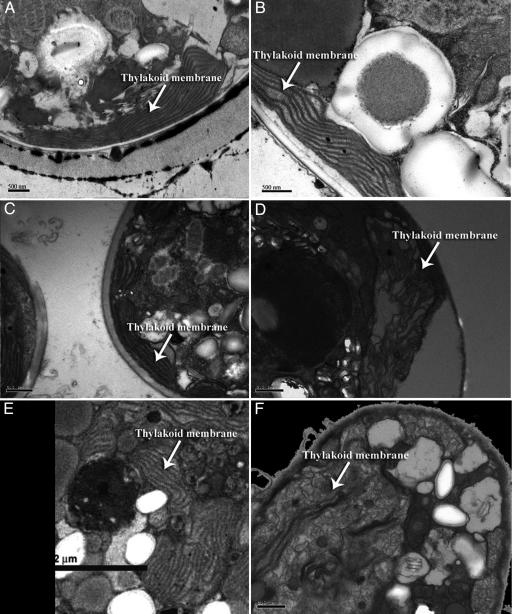
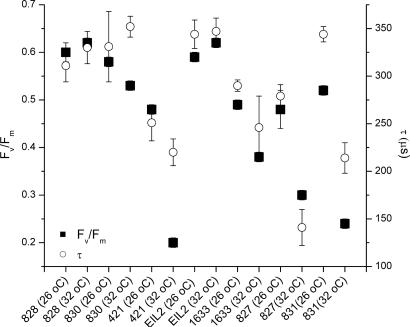
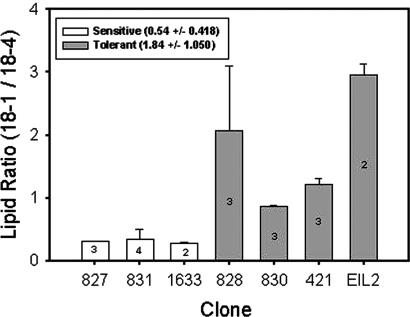
Over the past three decades, massive bleaching events of zooxanthellate corals have been documented across the range of global distribution. Although the phenomenon is correlated with relatively small increases in sea-surface temperature and enhanced light intensity, the underlying physiological mechanism remains unknown. In this article we demonstrate that thylakoid membrane lipid composition is a key determinate of thermal-stress sensitivity in symbiotic algae of cnidarians. Analyses of thylakoid membranes reveal that the critical threshold temperature separating thermally tolerant from sensitive species of zooxanthellae is determined by the saturation of the lipids. The lipid composition is potentially diagnostic of the differential nature of thermally induced bleaching found in scleractinian corals. Measurements of variable chlorophyll fluorescence kinetic transients indicate that thermally damaged membranes are energetically uncoupled but remain capable of splitting water. Consequently, a fraction of the photosynthetically produced oxygen is reduced by photosystem I through the Mehler reaction to form reactive oxygen species, which rapidly accumulate at high irradiance levels and trigger death and expulsion of the endosymbiotic algae. Differential sensitivity to thermal stress among the various species of Symbiodinium seems to be distributed across all clades. A clocked molecular phylogenetic analysis suggests that the evolutionary history of symbiotic algae in cnidarians selected for a reduced tolerance to elevated temperatures in the latter portion of the Cenozoic.
Figures
References
-
- Hughes, T. P., Baird, A. H., Bellwood, D. R., Card, M., Connolly, S. R., Folke, C., Grosberg, R., Hoegh-Guldberg, O., Jackson, J. B., Kleypas, J., et al. (2003) Science 301, 929–933. - PubMed
-
- Wilkinson, C. (2000) Status of Coral Reefs of the World: 2000 (Global Coral Reef Monitoring Network and Australian Institute of Marine Science, Townsville, Australia).
-
- Podesta, G. P. & Glynn, P. W. (2001) Bull. Mar. Sci. 69, 43–59.
-
- Baker, A. C. (2003) Annu. Rev. Ecol. Evol. Syst. 34, 661–689.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
