

Tumor-promoting phorbol esters and activated Ras inactivate the tuberous sclerosis tumor suppressor complex via p90 ribosomal S6 kinase
- PMID: 15342917
- PMCID: PMC518784
- DOI: 10.1073/pnas.0405659101
Tumor-promoting phorbol esters and activated Ras inactivate the tuberous sclerosis tumor suppressor complex via p90 ribosomal S6 kinase
Abstract
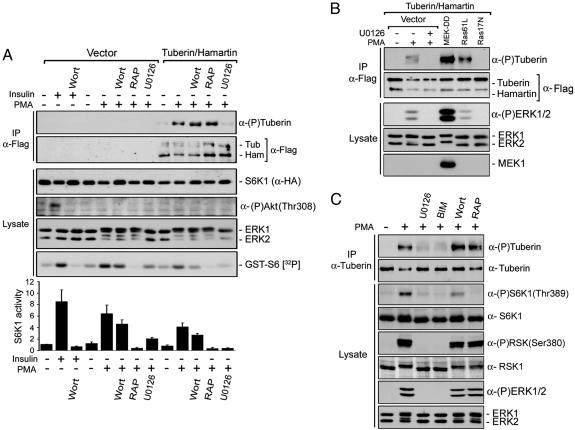
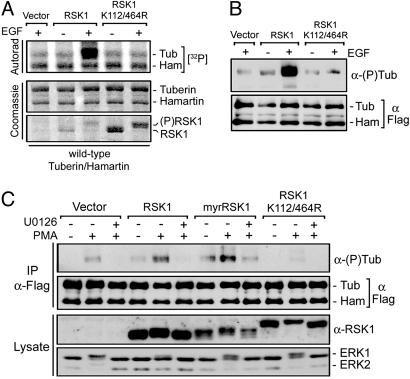
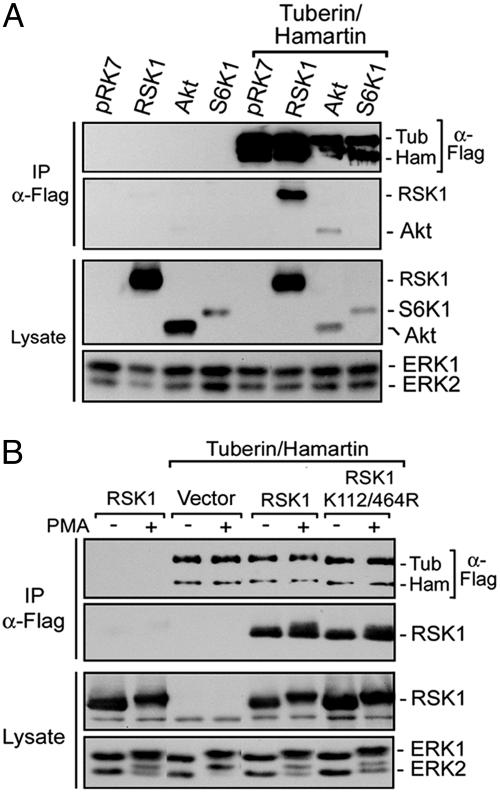
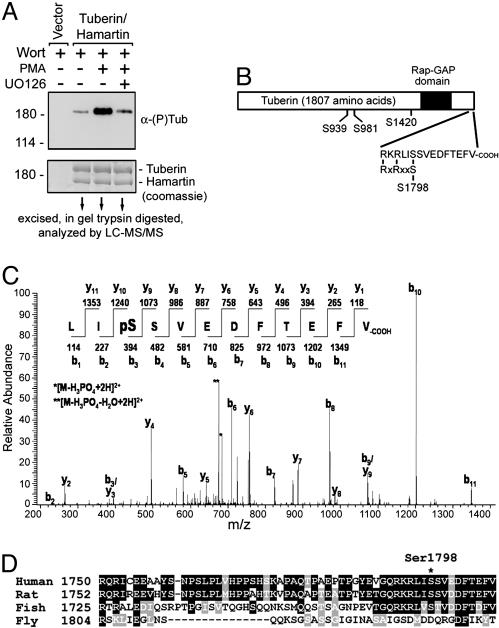
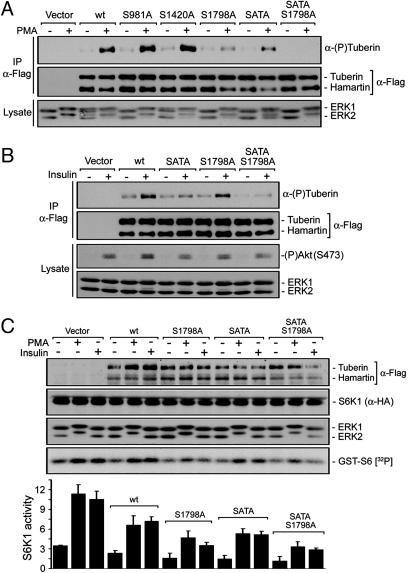
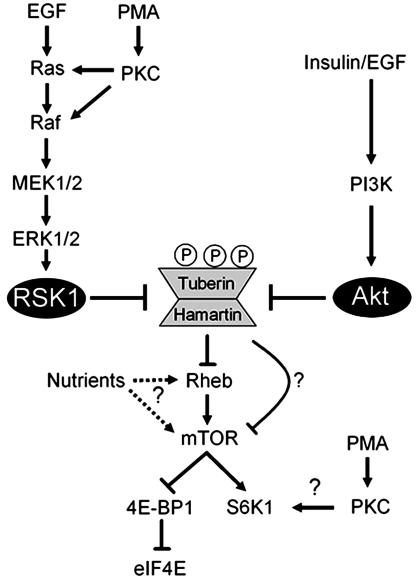
Tuberous sclerosis complex (TSC) is a genetic disorder caused by mutations in either of the two tumor suppressor genes TSC1 or TSC2, which encode hamartin and tuberin, respectively. Tuberin and hamartin form a complex that inhibits signaling by the mammalian target of rapamycin (mTOR), a critical nutrient sensor and regulator of cell growth and proliferation. Phosphatidylinositol 3-kinase (PI3K) inactivates the tumor suppressor complex and enhances mTOR signaling by means of phosphorylation of tuberin by Akt. Importantly, cellular transformation mediated by phorbol esters and Ras isoforms that poorly activate PI3K promote tumorigenesis in the absence of Akt activation. In this study, we show that phorbol esters and activated Ras also induce the phosphorylation of tuberin and collaborates with the nutrient-sensing pathway to regulate mTOR effectors, such as p70 ribosomal S6 kinase 1 (S6K1). The mitogen-activated protein kinase (MAPK)-activated kinase, p90 ribosomal S6 kinase (RSK) 1, was found to interact with and phosphorylate tuberin at a regulatory site, Ser-1798, located at the evolutionarily conserved C terminus of tuberin. RSK1 phosphorylation of Ser-1798 inhibits the tumor suppressor function of the tuberin/hamartin complex, resulting in increased mTOR signaling to S6K1. Together, our data unveil a regulatory mechanism by which the Ras/MAPK and PI3K pathways converge on the tumor suppressor tuberin to inhibit its function.
Figures
References
-
- Cheadle, J. P., Reeve, M. P., Sampson, J. R. & Kwiatkowski, D. J. (2000) Hum. Genet. 107, 97–114. - PubMed
-
- Carbonara, C., Longa, L., Grosso, E., Borrone, C., Garre, M. G., Brisigotti, M. & Migone, N. (1994) Hum. Mol. Genet. 3, 1829–1832. - PubMed
-
- Green, A. J., Smith, M. & Yates, J. R. (1994) Nat. Genet. 6, 193–196. - PubMed
-
- Ito, N. & Rubin, G. M. (1999) Cell 96, 529–539. - PubMed
-
- van Slegtenhorst, M., Nellist, M., Nagelkerken, B., Cheadle, J., Snell, R., van den Ouweland, A., Reuser, A., Sampson, J., Halley, D. & van der Sluijs, P. (1998) Hum. Mol. Genet. 7, 1053–1057. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
