

Adaptive evolution of centromere proteins in plants and animals
- PMID: 15345035
- PMCID: PMC549713
- DOI: 10.1186/jbiol11
Adaptive evolution of centromere proteins in plants and animals
Abstract
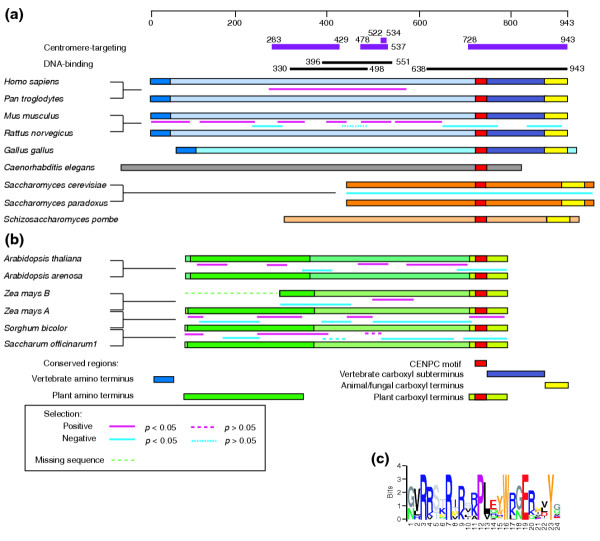
Background: Centromeres represent the last frontiers of plant and animal genomics. Although they perform a conserved function in chromosome segregation, centromeres are typically composed of repetitive satellite sequences that are rapidly evolving. The nucleosomes of centromeres are characterized by a special H3-like histone (CenH3), which evolves rapidly and adaptively in Drosophila and Arabidopsis. Most plant, animal and fungal centromeres also bind a large protein, centromere protein C (CENP-C), that is characterized by a single 24 amino-acid motif (CENPC motif).
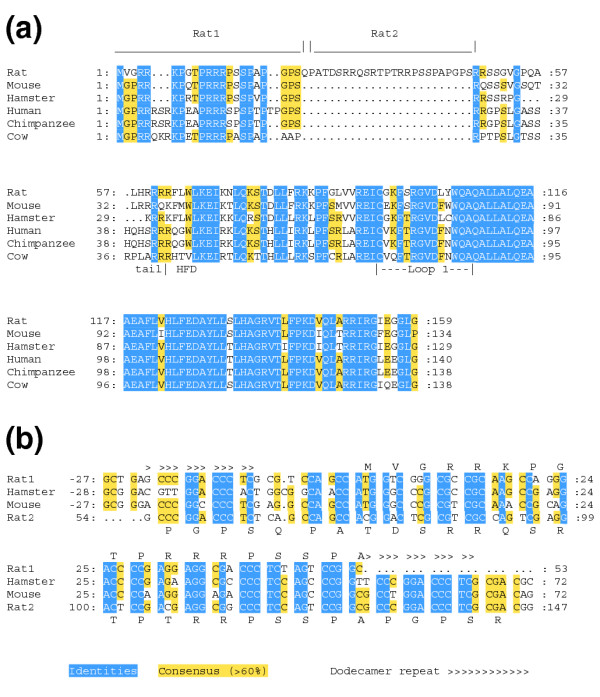
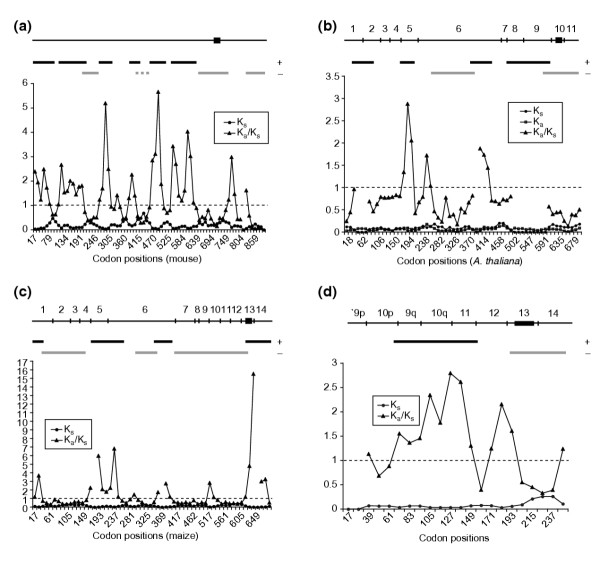
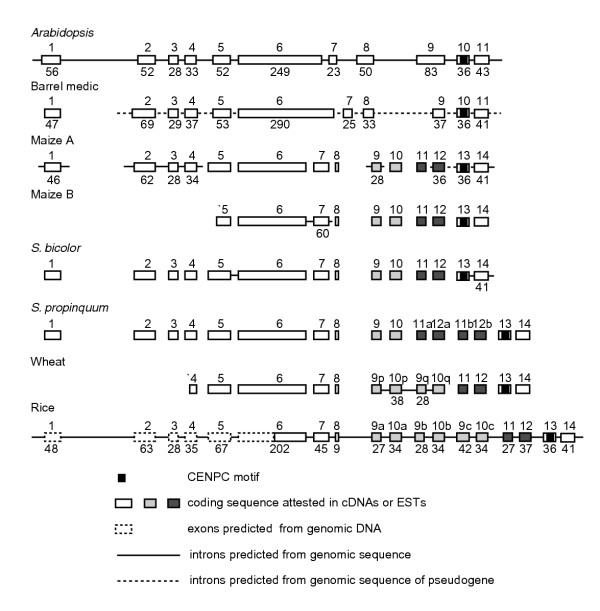
Results: Whereas we find no evidence that mammalian CenH3 (CENP-A) has been evolving adaptively, mammalian CENP-C proteins contain adaptively evolving regions that overlap with regions of DNA-binding activity. In plants we find that CENP-C proteins have complex duplicated regions, with conserved amino and carboxyl termini that are dissimilar in sequence to their counterparts in animals and fungi. Comparisons of Cenpc genes from Arabidopsis species and from grasses revealed multiple regions that are under positive selection, including duplicated exons in some grasses. In contrast to plants and animals, yeast CENP-C (Mif2p) is under negative selection.
Conclusions: CENP-Cs in all plant and animal lineages examined have regions that are rapidly and adaptively evolving. To explain these remarkable evolutionary features for a single-copy gene that is needed at every mitosis, we propose that CENP-Cs, like some CenH3s, suppress meiotic drive of centromeres during female meiosis. This process can account for the rapid evolution and the complexity of centromeric DNA in plants and animals as compared to fungi.
Figures

References
-
- Lohe A, Roberts P. Evolution of satellite DNA sequences in Drosophila. In: Verma RS, editor. In Heterochromatin, Molecular and Structural Aspects. Cambridge: Cambridge University Press; 1988. pp. 148–186.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
