

Discovery of estrogen receptor alpha target genes and response elements in breast tumor cells
- PMID: 15345050
- PMCID: PMC522873
- DOI: 10.1186/gb-2004-5-9-r66
Discovery of estrogen receptor alpha target genes and response elements in breast tumor cells
Abstract
Background: Estrogens and their receptors are important in human development, physiology and disease. In this study, we utilized an integrated genome-wide molecular and computational approach to characterize the interaction between the activated estrogen receptor (ER) and the regulatory elements of candidate target genes.
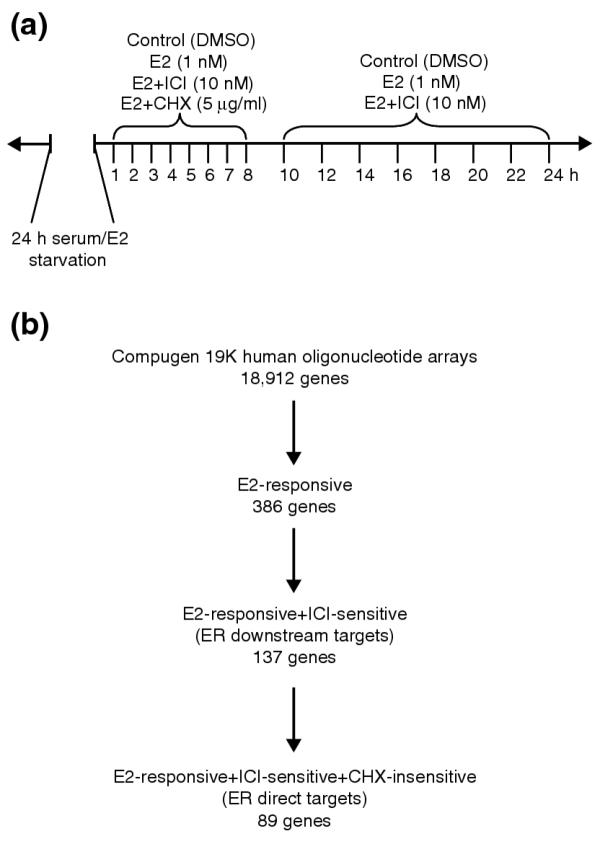
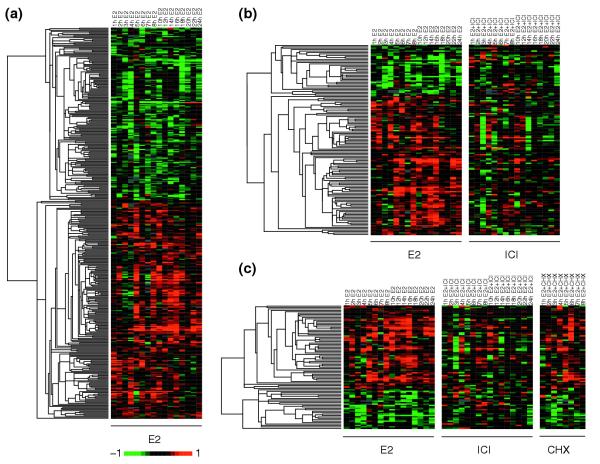
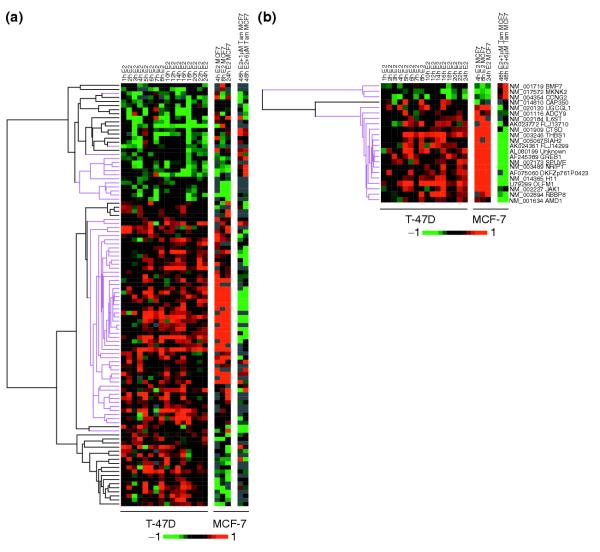
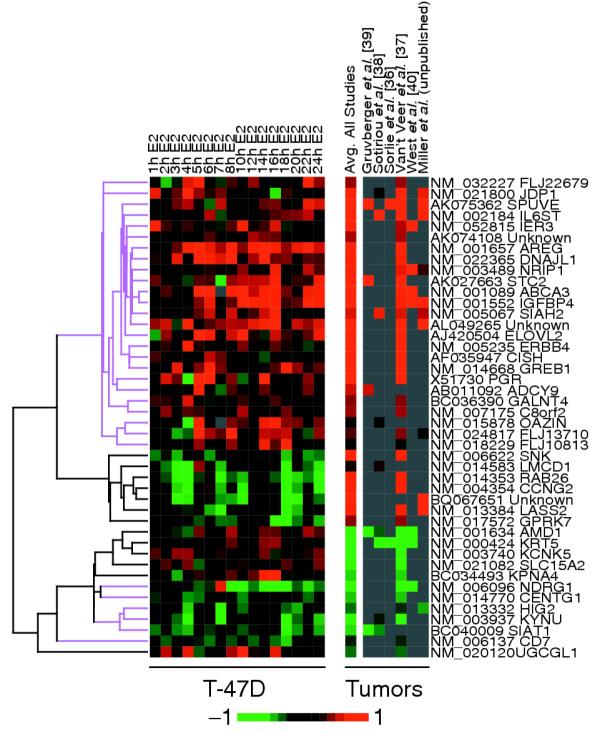
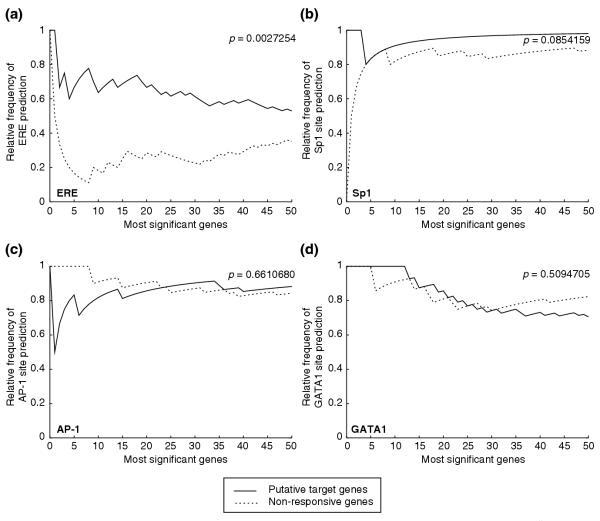
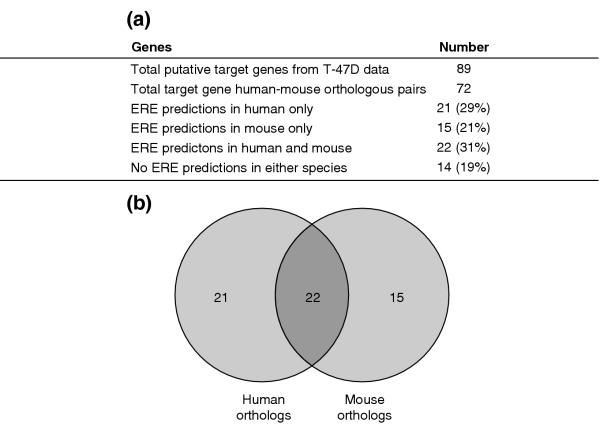
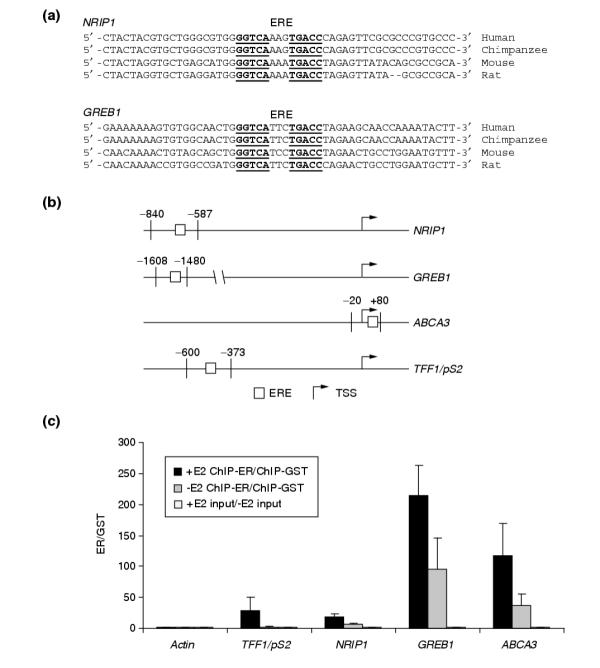
Results: Of around 19,000 genes surveyed in this study, we observed 137 ER-regulated genes in T-47D cells, of which only 89 were direct target genes. Meta-analysis of heterogeneous in vitro and in vivo datasets showed that the expression profiles in T-47D and MCF-7 cells are remarkably similar and overlap with genes differentially expressed between ER-positive and ER-negative tumors. Computational analysis revealed a significant enrichment of putative estrogen response elements (EREs) in the cis-regulatory regions of direct target genes. Chromatin immunoprecipitation confirmed ligand-dependent ER binding at the computationally predicted EREs in our highest ranked ER direct target genes, NRIP1, GREB1 and ABCA3. Wider examination of the cis-regulatory regions flanking the transcriptional start sites showed species conservation in mouse-human comparisons in only 6% of predicted EREs.
Conclusions: Only a small core set of human genes, validated across experimental systems and closely associated with ER status in breast tumors, appear to be sufficient to induce ER effects in breast cancer cells. That cis-regulatory regions of these core ER target genes are poorly conserved suggests that different evolutionary mechanisms are operative at transcriptional control elements than at coding regions. These results predict that certain biological effects of estrogen signaling will differ between mouse and human to a larger extent than previously thought.
Figures
References
-
- Nilsson S, Gustafsson JÅ. Estrogen receptor action. Crit Rev Eukaryot Gene Expr. 2002;12:237–257. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
