

Organization of Ca2+ release units in excitable smooth muscle of the guinea-pig urinary bladder
- PMID: 15345562
- PMCID: PMC1304588
- DOI: 10.1529/biophysj.104.044123
Organization of Ca2+ release units in excitable smooth muscle of the guinea-pig urinary bladder
Erratum in
- Biophys J. 2004 Oct;87(4):2914
Abstract
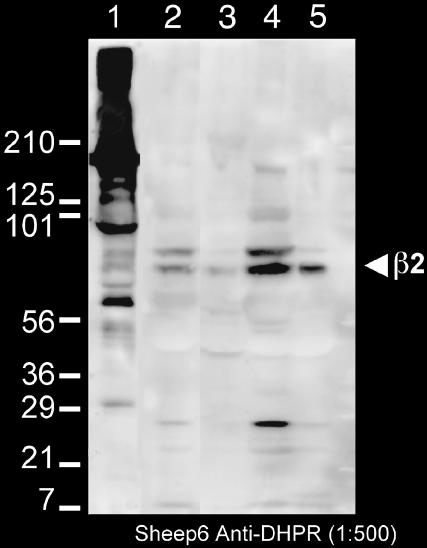
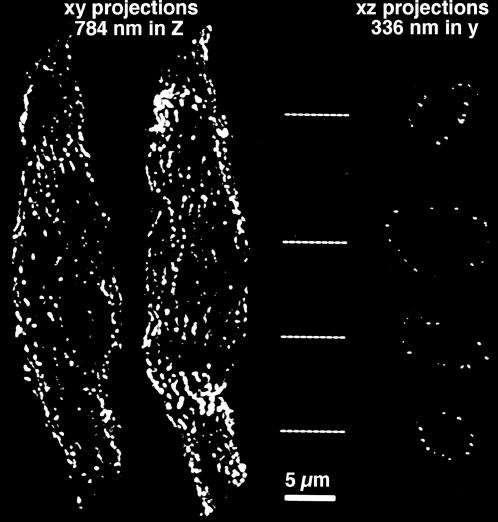
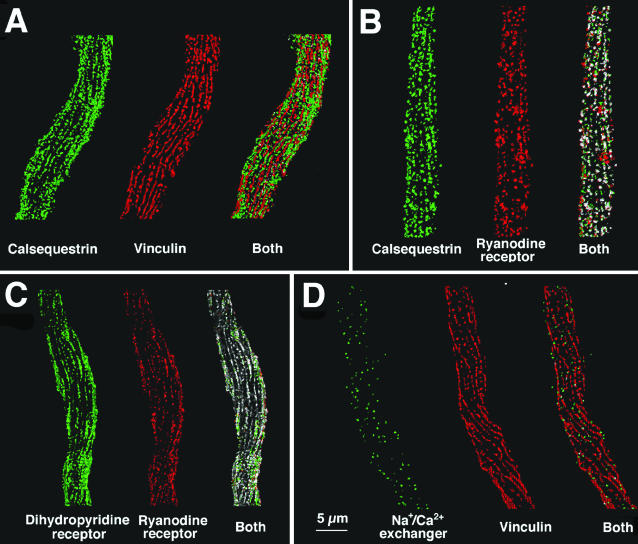
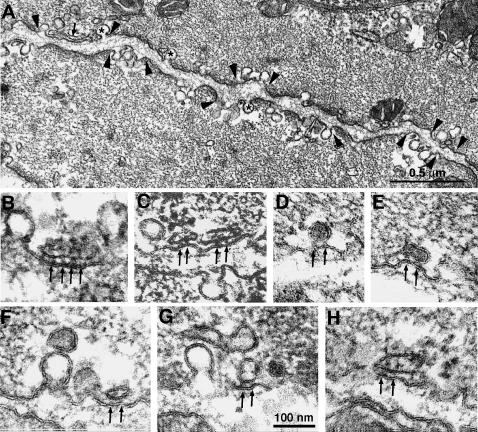
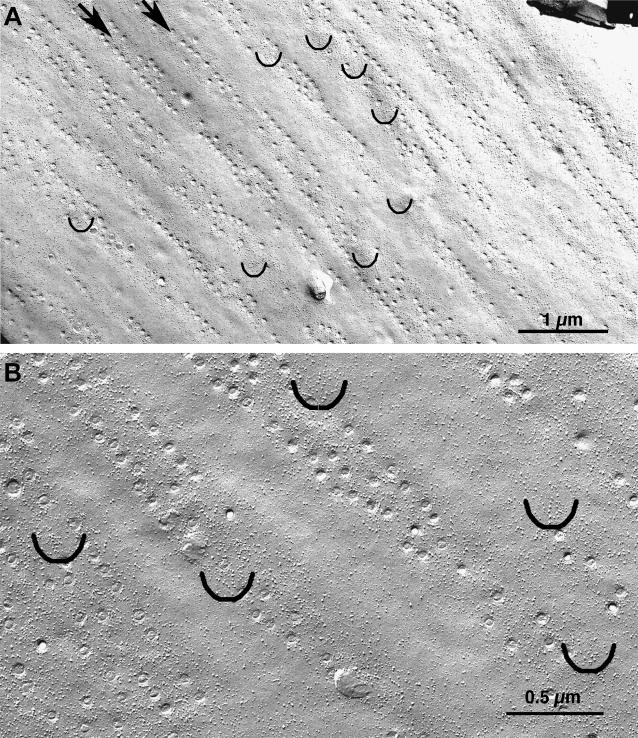
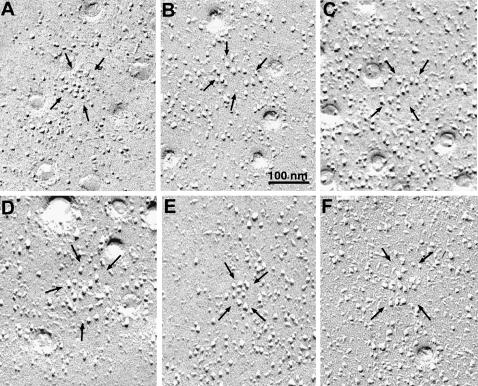
Ca(2+) release from internal stores (sarcoplasmic reticulum or SR) in smooth muscles is initiated either via pharmaco-mechanical coupling due to the action of an agonist and involving IP3 receptors, or via excitation-contraction coupling, mostly involving L-type calcium channels in the plasmalemma (DHPRs), and ryanodine receptors (RyRs), or Ca(2+) release channels of the SR. This work focuses attention on the structural basis for the coupling between DHPRs and RyRs in phasic smooth muscle cells of the guinea-pig urinary bladder. Immunolabeling shows that two proteins of the SR: calsequestrin and the RyR, and one protein the plasmalemma, the L-type channel or DHPR, are colocalized with each other within numerous, peripherally located sites located within the caveolar domains. Electron microscopy images from thin sections and freeze-fracture replicas identify feet in small peripherally located SR vesicles containing calsequestrin and distinctive large particles clustered within small membrane areas. Both feet and particle clusters are located within caveolar domains. Correspondence between the location of feet and particle clusters and of RyR- and DHPR-positive foci allows the conclusion that calsequestrin, RyRs, and L-type Ca(2+) channels are associated with peripheral couplings, or Ca(2+) release units, constituting the key machinery involved in excitation-contraction coupling. Structural analogies between smooth and cardiac muscle excitation-contraction coupling complexes suggest a common basic mechanism of action.
Figures
References
-
- Arikkath, J., C. C. Chen, C. Ahern, V. Allamand, J. D. Flanagan, R. Coronado, R. G. Gregg, and K. P. Campbell. 2003. Gamma 1 subunit interactions within the skeletal muscle L-type voltage-gated calcium channels. J. Biol. Chem. 278:1212–1219. - PubMed
-
- Bolton, T. B., and D. V. Gordienko. 1998. Confocal imaging of calcium release events in single smooth muscle cells. Acta Physiol. Scand. 164:567–575. - PubMed
-
- Bolton, T. B., S. A. Prestwic, A. V. Zholos, and D. V. Gordienko. 1999. Excitation-contraction coupling in gastrointestinal and other smooth muscles. Annu. Rev. Physiol. 61:85–115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
