

Down-regulation of DELLA genes is not essential for germination of tomato, soybean, and Arabidopsis seeds
- PMID: 15347801
- PMCID: PMC523341
- DOI: 10.1104/pp.103.034876
Down-regulation of DELLA genes is not essential for germination of tomato, soybean, and Arabidopsis seeds
Abstract
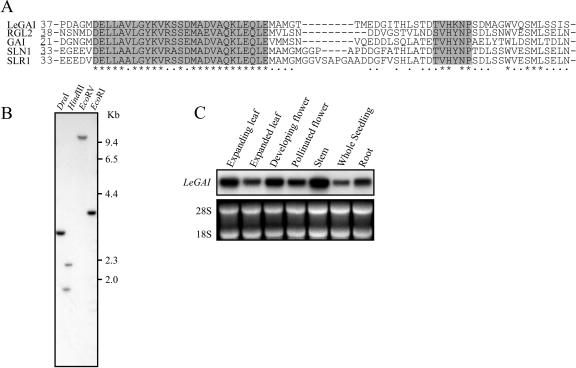
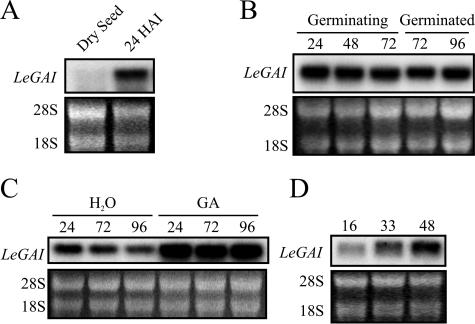
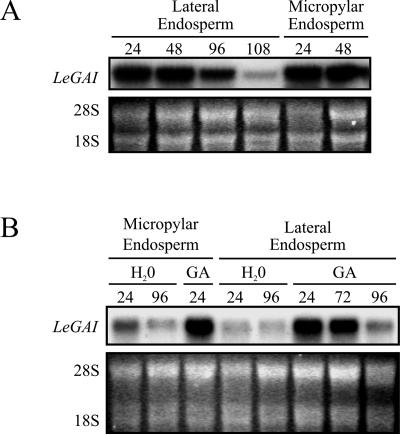
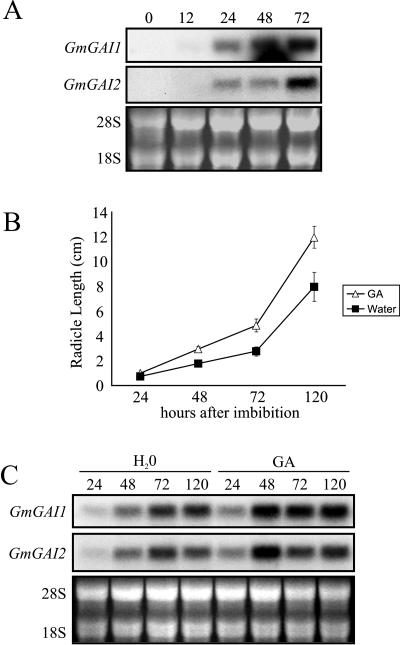
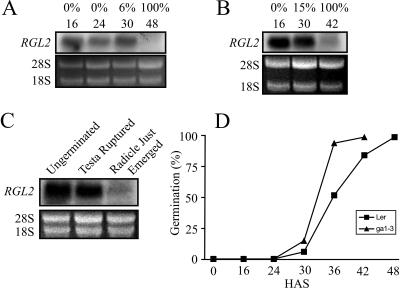
The relationship between expression of a negative regulator of GA signal transduction (RGL2) belonging to the DELLA gene family and repression of Arabidopsis seed germination has been studied (Lee S, Cheng H, King KE, Wang W, He Y, Hussain A, Lo J, Harberd NP, Peng J [2002] Genes and Development 16: 646-658). There is one DELLA gene (LeGAI) present in tomato (Lycopersicon esculentum Mill.), which is expressed in both vegetative and reproductive tissues. During germination of wild-type tomato seed, there was no decline in the expression of LeGAI in either the embryo or the endosperm. Rather, LeGAI transcripts increased in these tissues following imbibition and remained high during and following germination. A similar increase in LeGAI transcripts occurred in the endosperm and embryo of GA-treated gib-1 mutant seed during and following germination. Likewise in soybean (Glycine max) seed, there was no decline in the expression of two DELLA genes in the radicle before or after germination. Upon reexamination of RGL2 in Arabidopsis seeds, a decline in its expression was noted but only after radicle emergence, i.e. after germination had been completed. Taken together, these data are consistent with GA-induced down-regulation of DELLA genes not being a prerequisite for germination of tomato, soybean, and Arabidopsis seeds.
Figures
References
-
- Bewley JD (1997. b) Breaking down the walls—a role for endo-β–mannanase in release from seed dormancy? Trends Plant Sci 2: 464–469
-
- Bewley JD, Black M (1994) Seeds: Physiology of Development and Germination, Ed 2. Plenum Press, New York
-
- Bradford KJ, Chen F, Cooley MB, Dahal P, Downie B, Fukunaga KK, Gee OH, Gurusinghe S, Mella RA, Nonogaki H, et al (2000) Gene expression prior to radicle emergence in imbibed tomato seeds. In M Black, KJ Bradford, J Vazquez-Ramos, eds, Seed Biology: Advances and Applications. CABI Publishing, Wallingford, UK, pp 231–251
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
