

Conserved extracellular cysteine residues and cytoplasmic loop-loop interplay are required for functionality of the heptahelical MLO protein
- PMID: 15352871
- PMCID: PMC1134693
- DOI: 10.1042/BJ20040993
Conserved extracellular cysteine residues and cytoplasmic loop-loop interplay are required for functionality of the heptahelical MLO protein
Abstract
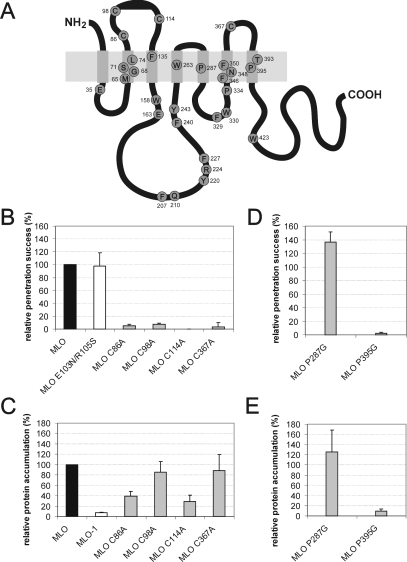
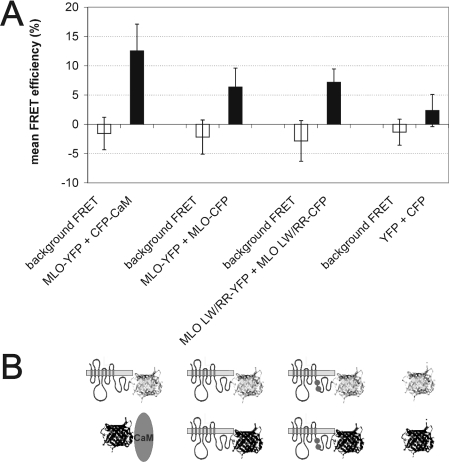
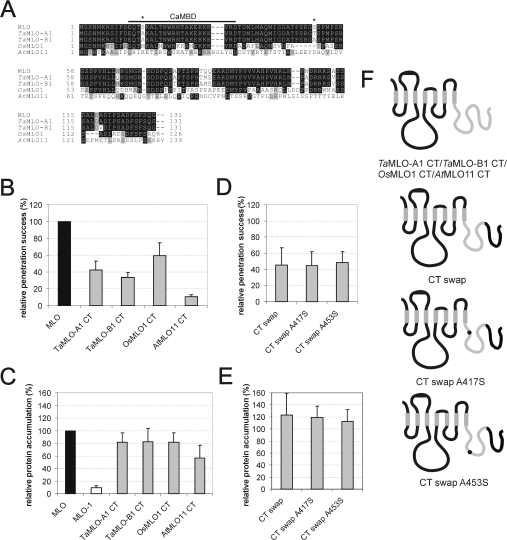
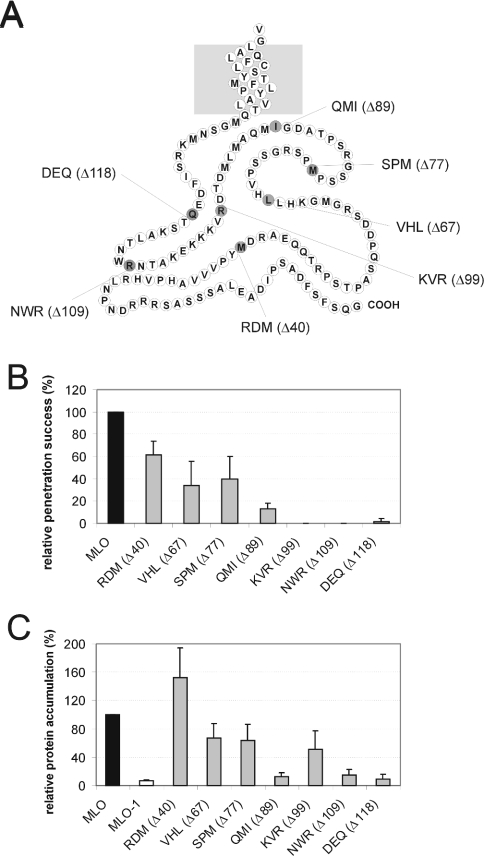
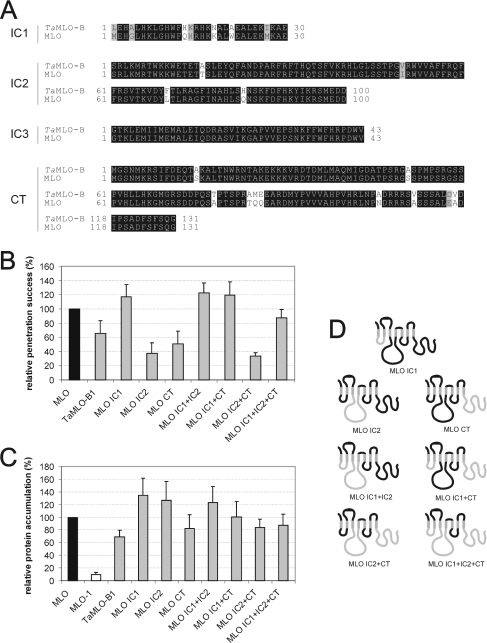
We performed a structure-function analysis of the plasma membrane-localized plant-specific barley (Hordeum vulgare) MLO (powdery-mildew-resistance gene o) protein. Invariant cysteine and proline residues, located either in extracellular loops or transmembrane domains that have been conserved in MLO proteins for more than 400 million years, were found to be essential for MLO functionality and/or stability. Similarly to many metazoan G-protein-coupled receptors known to function as homo- and hetero-oligomers, FRET (fluorescence resonance energy transfer) analysis revealed evidence for in planta MLO dimerization/oligomerization. Domain-swap experiments with closely related wheat and rice as well as diverged Arabidopsis MLO isoforms demonstrated that the identity of the C-terminal cytoplasmic tail contributes to MLO activity. Likewise, analysis of a progressive deletion series revealed that integrity of the C-terminus determines both MLO accumulation and functionality. A series of domain swaps of cytoplasmic loops with the wheat (Triticum aestivum) orthologue, TaMLO-B1, provided strong evidence for co-operative loop-loop interplay either within the protein or between MLO molecules. Our data indicate extensive intramolecular co-evolution of cytoplasmic domains in the evolutionary history of the MLO protein family.
Figures
Similar articles
-
Functional conservation of wheat and rice Mlo orthologs in defense modulation to the powdery mildew fungus.Mol Plant Microbe Interact. 2002 Oct;15(10):1069-77. doi: 10.1094/MPMI.2002.15.10.1069. Mol Plant Microbe Interact. 2002. PMID: 12437305
-
Novel induced mlo mutant alleles in combination with site-directed mutagenesis reveal functionally important domains in the heptahelical barley Mlo protein.BMC Plant Biol. 2010 Feb 19;10:31. doi: 10.1186/1471-2229-10-31. BMC Plant Biol. 2010. PMID: 20170486 Free PMC article.
-
Topology, subcellular localization, and sequence diversity of the Mlo family in plants.J Biol Chem. 1999 Dec 3;274(49):34993-5004. doi: 10.1074/jbc.274.49.34993. J Biol Chem. 1999. PMID: 10574976
-
Serpentine plant MLO proteins as entry portals for powdery mildew fungi.Biochem Soc Trans. 2005 Apr;33(Pt 2):389-92. doi: 10.1042/BST0330389. Biochem Soc Trans. 2005. PMID: 15787613 Review.
-
Functional domains in tetraspanin proteins.Trends Biochem Sci. 2003 Feb;28(2):106-12. doi: 10.1016/S0968-0004(02)00014-2. Trends Biochem Sci. 2003. PMID: 12575999 Review.
Cited by
-
Genome-wide characterization and expression analysis of the MLO gene family sheds light on powdery mildew resistance in Lagenaria siceraria.Heliyon. 2023 Mar 24;9(4):e14624. doi: 10.1016/j.heliyon.2023.e14624. eCollection 2023 Apr. Heliyon. 2023. PMID: 37025859 Free PMC article.
-
Monocot and dicot MLO powdery mildew susceptibility factors are functionally conserved in spite of the evolution of class-specific molecular features.BMC Plant Biol. 2015 Oct 26;15:257. doi: 10.1186/s12870-015-0639-6. BMC Plant Biol. 2015. PMID: 26499889 Free PMC article.
-
MLO Differentially Regulates Barley Root Colonization by Beneficial Endophytic and Mycorrhizal Fungi.Front Plant Sci. 2020 Jan 16;10:1678. doi: 10.3389/fpls.2019.01678. eCollection 2019. Front Plant Sci. 2020. PMID: 32010163 Free PMC article.
-
Comparative phylogenetic analysis of genome-wide Mlo gene family members from Glycine max and Arabidopsis thaliana.Mol Genet Genomics. 2014 Jun;289(3):345-59. doi: 10.1007/s00438-014-0811-y. Epub 2014 Jan 28. Mol Genet Genomics. 2014. PMID: 24469270
-
The visible touch: in planta visualization of protein-protein interactions by fluorophore-based methods.Plant Methods. 2006 Jun 26;2:12. doi: 10.1186/1746-4811-2-12. Plant Methods. 2006. PMID: 16800872 Free PMC article.
References
-
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature (London) 2000;408:796–815. - PubMed
-
- Goff S. A., Ricke D., Lan T. H., Presting G., Wang R. L., Dunn M., Glazebrook J., Sessions A., Oeller P., Varma H., et al. A draft sequence of the rice genome (Oryza sativa L. ssp japonica) Science. 2002;296:92–100. - PubMed
-
- Yu J., Hu S. N., Wang J., Wong G. K. S., Li S. G., Liu B., Deng Y. J., Dai L., Zhou Y., Zhang X. Q., et al. A draft sequence of the rice genome (Oryza sativa L. ssp indica) Science. 2002;296:79–92. - PubMed
-
- Büschges R., Hollricher K., Panstruga R., Simons G., Wolter M., Frijters A., van Daelen R., van der Lee T., Diergaarde P., Groenendijk J., et al. The barley Mlo gene: a novel control element of plant pathogen resistance. Cell. 1997;88:695–705. - PubMed
-
- Devoto A., Piffanelli P., Nilsson I., Wallin E., Panstruga R., von Heijne G., Schulze-Lefert P. Topology, subcellular localization, and sequence diversity of the Mlo family in plants. J. Biol. Chem. 1999;274:34993–35004. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases