

Visualization of early chromosome condensation: a hierarchical folding, axial glue model of chromosome structure
- PMID: 15353545
- PMCID: PMC2172117
- DOI: 10.1083/jcb.200406049
Visualization of early chromosome condensation: a hierarchical folding, axial glue model of chromosome structure
Abstract
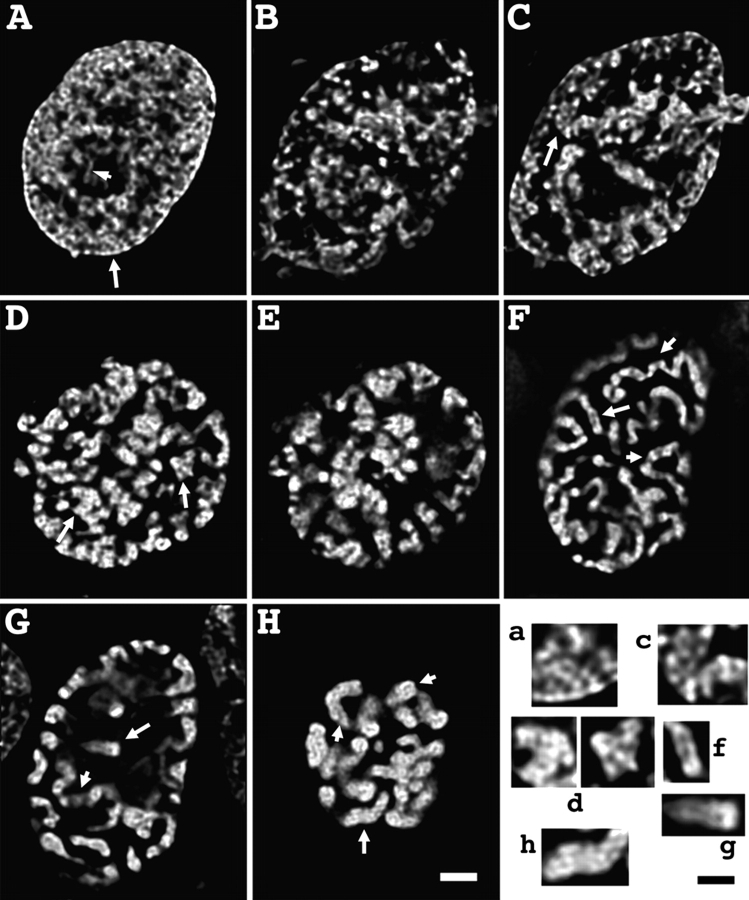
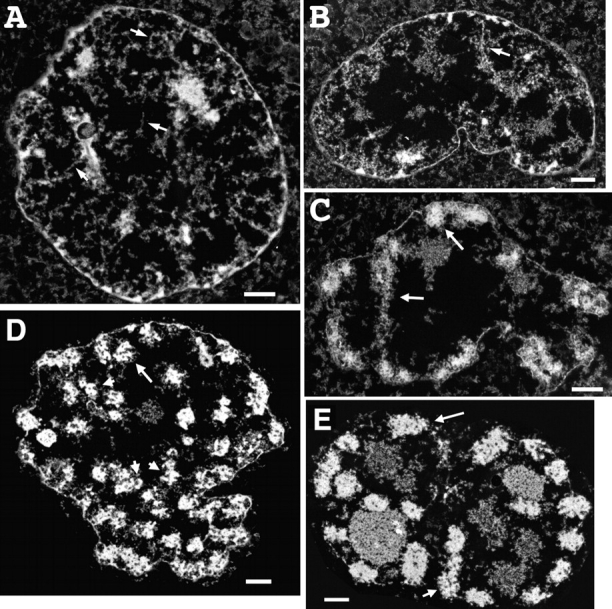
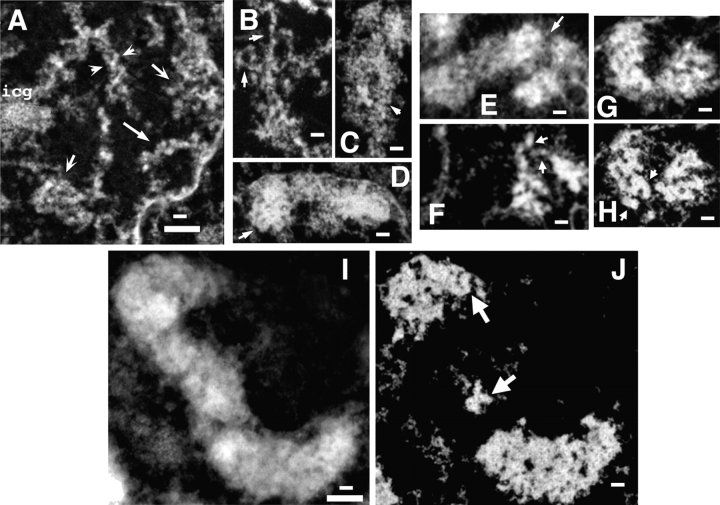
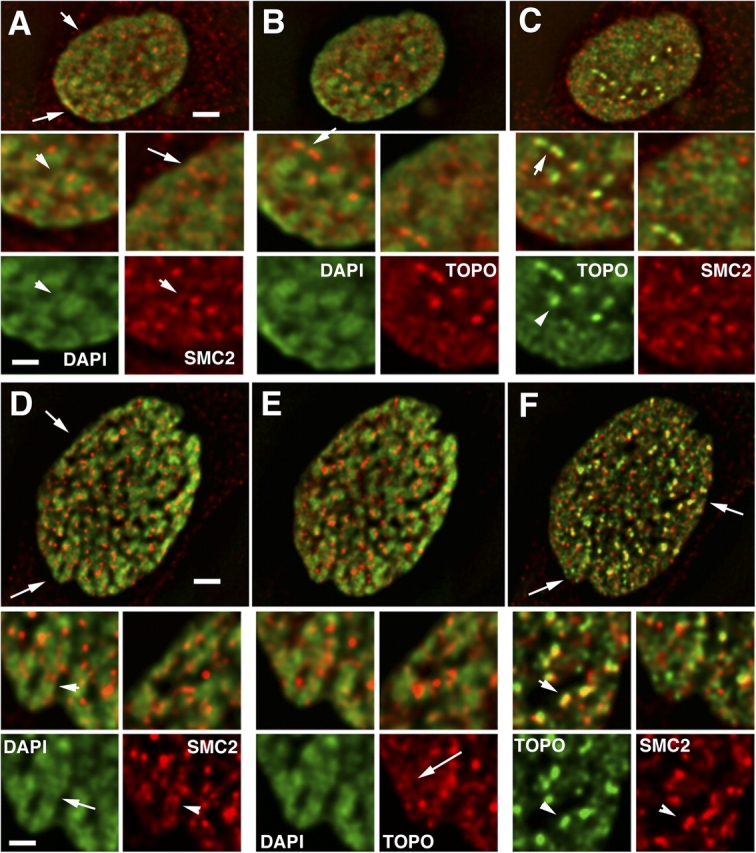
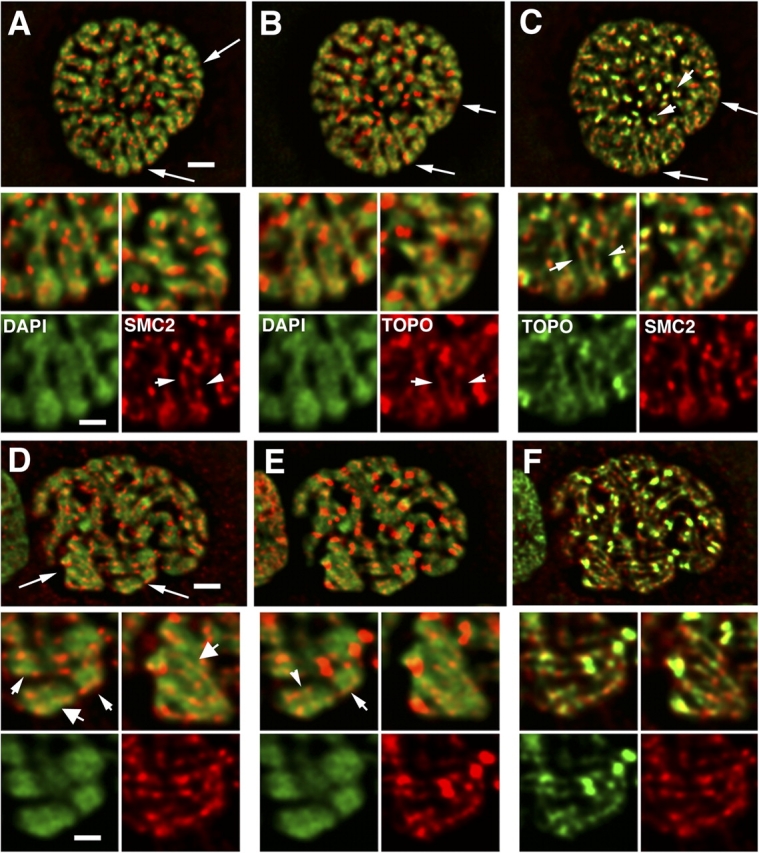
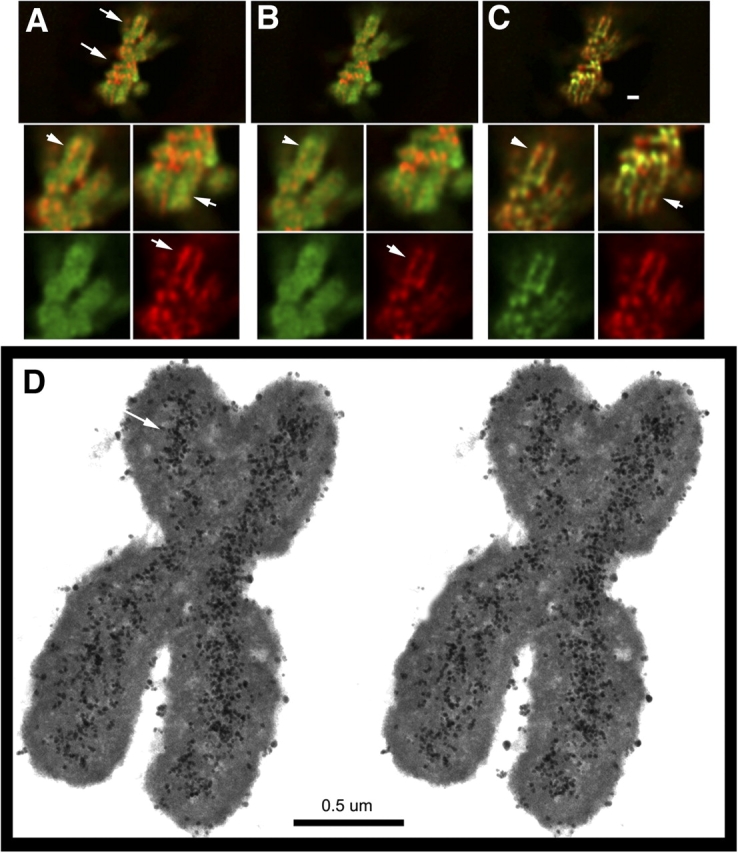
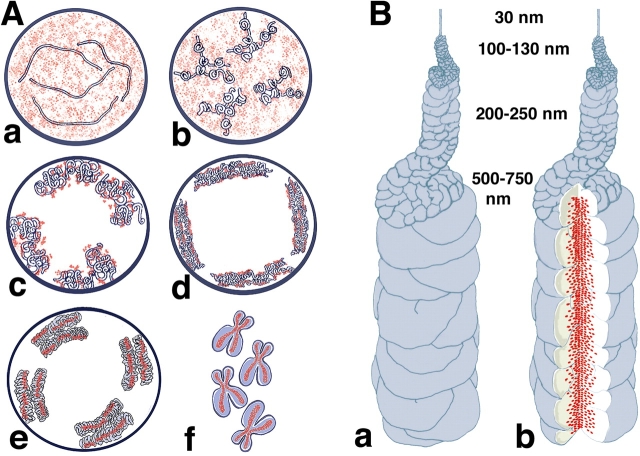
Current models of mitotic chromosome structure are based largely on the examination of maximally condensed metaphase chromosomes. Here, we test these models by correlating the distribution of two scaffold components with the appearance of prophase chromosome folding intermediates. We confirm an axial distribution of topoisomerase IIalpha and the condensin subunit, structural maintenance of chromosomes 2 (SMC2), in unextracted metaphase chromosomes, with SMC2 localizing to a 150-200-nm-diameter central core. In contrast to predictions of radial loop/scaffold models, this axial distribution does not appear until late prophase, after formation of uniformly condensed middle prophase chromosomes. Instead, SMC2 associates throughout early and middle prophase chromatids, frequently forming foci over the chromosome exterior. Early prophase condensation occurs through folding of large-scale chromatin fibers into condensed masses. These resolve into linear, 200-300-nm-diameter middle prophase chromatids that double in diameter by late prophase. We propose a unified model of chromosome structure in which hierarchical levels of chromatin folding are stabilized late in mitosis by an axial "glue."
Figures
References
-
- Belmont, A.S. 1998. Nuclear ultrastructure: transmission electron microscopy and image analysis. Methods Cell Biol. 53:99–124. - PubMed
-
- Boy de la Tour, E., and U.K. Laemmli. 1988. The metaphase scaffold is helically folded: sister chromatids have predominately opposite helical handedness. Cell. 55:937–944. - PubMed
