

Bacterial transcriptional regulators for degradation pathways of aromatic compounds
- PMID: 15353566
- PMCID: PMC515250
- DOI: 10.1128/MMBR.68.3.474-500.2004
Bacterial transcriptional regulators for degradation pathways of aromatic compounds
Abstract
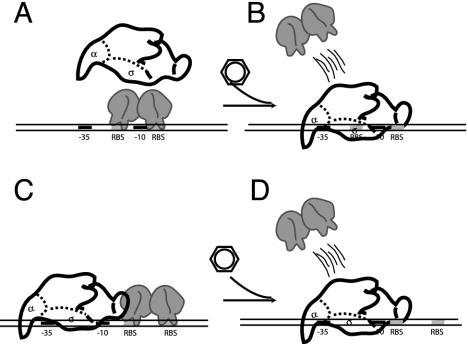
Human activities have resulted in the release and introduction into the environment of a plethora of aromatic chemicals. The interest in discovering how bacteria are dealing with hazardous environmental pollutants has driven a large research community and has resulted in important biochemical, genetic, and physiological knowledge about the degradation capacities of microorganisms and their application in bioremediation, green chemistry, or production of pharmacy synthons. In addition, regulation of catabolic pathway expression has attracted the interest of numerous different groups, and several catabolic pathway regulators have been exemplary for understanding transcription control mechanisms. More recently, information about regulatory systems has been used to construct whole-cell living bioreporters that are used to measure the quality of the aqueous, soil, and air environment. The topic of biodegradation is relatively coherent, and this review presents a coherent overview of the regulatory systems involved in the transcriptional control of catabolic pathways. This review summarizes the different regulatory systems involved in biodegradation pathways of aromatic compounds linking them to other known protein families. Specific attention has been paid to describing the genetic organization of the regulatory genes, promoters, and target operon(s) and to discussing present knowledge about signaling molecules, DNA binding properties, and operator characteristics, and evidence from regulatory mutants. For each regulator family, this information is combined with recently obtained protein structural information to arrive at a possible mechanism of transcription activation. This demonstrates the diversity of control mechanisms existing in catabolic pathways.
Figures
Similar articles
-
Specific and global regulation of genes associated with the degradation of aromatic compounds in bacteria.J Mol Microbiol Biotechnol. 2002 Mar;4(2):111-21. J Mol Microbiol Biotechnol. 2002. PMID: 11873906 Review.
-
Metabolic reconstruction of aromatic compounds degradation from the genome of the amazing pollutant-degrading bacterium Cupriavidus necator JMP134.FEMS Microbiol Rev. 2008 Aug;32(5):736-94. doi: 10.1111/j.1574-6976.2008.00122.x. Epub 2008 Aug 7. FEMS Microbiol Rev. 2008. PMID: 18691224 Review.
-
Transcriptional regulation of catabolic pathways for aromatic compounds in Corynebacterium glutamicum.Genet Mol Res. 2006 Dec 7;5(4):773-89. Genet Mol Res. 2006. PMID: 17183485 Review.
-
Regulation of genes in Streptomyces bacteria required for catabolism of lignin-derived aromatic compounds.Appl Microbiol Biotechnol. 2010 Apr;86(3):921-9. doi: 10.1007/s00253-009-2358-0. Epub 2009 Dec 9. Appl Microbiol Biotechnol. 2010. PMID: 20012281
-
Degradation and assimilation of aromatic compounds by Corynebacterium glutamicum: another potential for applications for this bacterium?Appl Microbiol Biotechnol. 2012 Jul;95(1):77-89. doi: 10.1007/s00253-012-4139-4. Epub 2012 May 17. Appl Microbiol Biotechnol. 2012. PMID: 22588501 Review.
Cited by
-
The Pathway-Specific Regulator ClaR of Streptomyces clavuligerus Has a Global Effect on the Expression of Genes for Secondary Metabolism and Differentiation.Appl Environ Microbiol. 2015 Oct;81(19):6637-48. doi: 10.1128/AEM.00916-15. Epub 2015 Jul 17. Appl Environ Microbiol. 2015. PMID: 26187955 Free PMC article.
-
Decoding the genetic networks of environmental bacteria: regulatory moonlighting of the TOL system of Pseudomonas putida mt-2.ISME J. 2013 Feb;7(2):229-32. doi: 10.1038/ismej.2012.98. Epub 2012 Aug 16. ISME J. 2013. PMID: 22895160 Free PMC article. No abstract available.
-
Diversity, abundance, and consistency of microbial oxygenase expression and biodegradation in a shallow contaminated aquifer.Appl Environ Microbiol. 2009 Oct;75(20):6478-87. doi: 10.1128/AEM.01091-09. Epub 2009 Aug 21. Appl Environ Microbiol. 2009. PMID: 19700556 Free PMC article.
-
The Integrative Conjugative Element clc (ICEclc) of Pseudomonas aeruginosa JB2.Front Microbiol. 2018 Jul 11;9:1532. doi: 10.3389/fmicb.2018.01532. eCollection 2018. Front Microbiol. 2018. PMID: 30050515 Free PMC article.
-
Ethylphenol Formation by Lactobacillus plantarum: Identification of the Enzyme Involved in the Reduction of Vinylphenols.Appl Environ Microbiol. 2018 Aug 17;84(17):e01064-18. doi: 10.1128/AEM.01064-18. Print 2018 Sep 1. Appl Environ Microbiol. 2018. PMID: 29934329 Free PMC article.
References
-
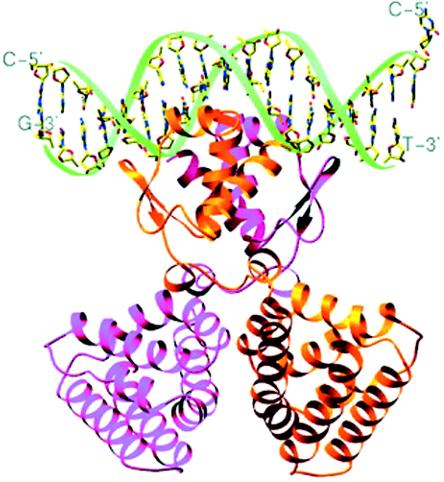
- Abril, M. A., M. Buck, and J. L. Ramos. 1991. Activation of the Pseudomonas TOL plasmid upper pathway operon. Identification of binding sites for the positive regulator XylR and for integration host factor protein. J. Biol. Chem. 266:15832-15838. - PubMed
-
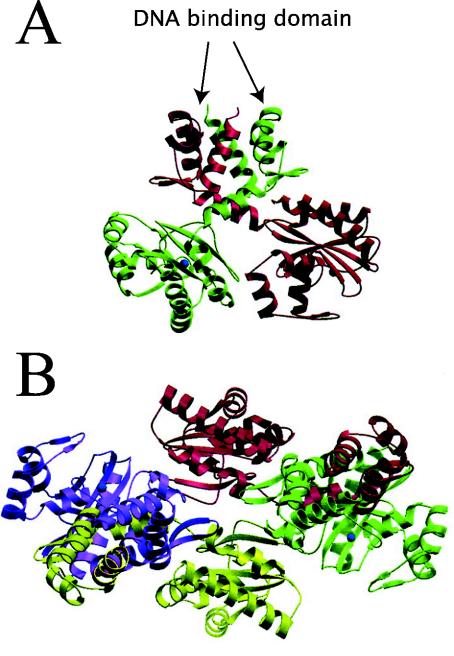
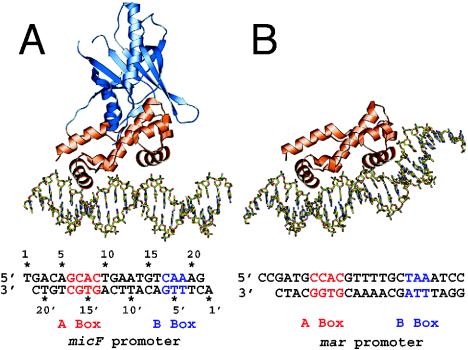
- Alekshun, M. N., S. B. Levy, T. R. Mealy, B. A. Seaton, and J. F. Head. 2001. The crystal structure of MarR, a regulator of multiple antibiotic resistance, at 2.3 Å resolution. Nat. Struct. Biol. 8:710-714. - PubMed
-
- Alex, L. A., and M. I. Simon. 1994. Protein histidine kinases and signal transduction in prokaryotes and eukaryotes. Trends Genet. 10:133-138. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
